![]()
PDF-Version
Neues aus der Forschung
Richard Lenski und die Evolution
Langzeitexperiment mit Bakterien belegt die Entstehung einer komplexen Neuerung

Ist die Evolution in der Lage, Innovationen hervorzubringen, die mehrere Organisationsschritte erfordern, bevor ein entsprechender Selektionsvorteil gegeben ist? Falls ja, welche Mechanismen leisten dies? Schaffen sie ihr Werk allmählich (über unzählige Generationen) oder sprunghaft (in einem einzelnen Organismus)? Zur Klärung dieser Fragen rief der US-amerikanische Evolutionsbiologe Richard LENSKI ein viel beachtetes Langzeitexperiment (LTEE) mit Bakterien ins Leben, das inzwischen seit über einem viertel Jahrhundert andauert. Tatsächlich hat die Studie bereits manch spektakuläre Erkenntnis geliefert.
Titelbild: Kompositbild des Langzeit-Evolutionsexperiments. Bildquelle: Richard Lenski und Brian Baer, Michigan State University. doi:10.1371/journal.pbio.1002185.g001, CC BY 4.0.
Bakterien als Forschungsobjekte
Bakterien haben gegenüber anderen Organismen den Vorteil, dass man sie in großer Zahl züchten kann. Zudem vermehren sie sich extrem rasch. Das macht sie zu idealen Forschungsobjekten, wenn man "Evolution in Aktion" sehen möchte.
Zu diesem Zweck rief ein Wissenschaftlerteam unter der Leitung des Evolutionsbiologen Richard LENSKI im Jahr 1988 das mittlerweile am längsten andauernde Evolutionsexperiment ins Leben. Studienobjekt war (und ist) das Bakterium Escherichia coli. Begonnen wurde mit einer einzigen Zelle, so dass alle Bakterien Nachfahren dieser einen Bakterienzelle sind. E. coli kann sich u.a. des Einfachzuckers Glucose als natürlicher Nahrungsquelle bedienen.
Zitronensäure macht den Unterschied
Im Gegensatz zur Glucose weiß der Wildtyp von E. coli nichts mit Citrat (Zitronensäure) als Kohlenstoffquelle anzufangen; der Phänotyp wird deshalb als Cit- bezeichnet. Zwar verstoffwechselt E. coli unter anaeroben Bedingungen, das heißt, unter Ausschluss von Sauerstoff, auch Citrat. An Luft (aerob) kann das Bakterium aus dieser Verbindung allerdings keine Energie gewinnen. Das Fehlen dieser Eigenschaft ist für E. coli derart spezifisch, dass es zur Unterscheidung von anderen Bakterienarten herangezogen wird.
Um zu prüfen, ob die Bakterien in der Lage sind, unter dem Druck starker Ressourcenknappheit dieser Beschränkung zu entkommen, übertrugen LENSKIs Kollegen zwei Klone des Wildtyps in ein Nährmedium. Dieses enthielt ein Minimalangebot an Glucose sowie ein Überangebot an nicht als Nahrungsquelle verwertbarem Citrat. Einer der Klone wies zudem eine "Knock-Out-Mutation" namens Ara- auf, die den Abbau des Zuckers Arabinose ausschaltet. Von beiden Klonen entnahmen die Wissenschaftler jeweils 6 Tochterzellen, die in frischem Nährmedium insgesamt 12 voneinander isolierte Starter-Kolonien bildeten.
Seit 37 Jahren (Stand: 2025) werden die Kolonien täglich aufgeteilt, wobei ein Anteil der Bakterien in neue Gefäße mit frischem Nährmedium übertragen werden, das meiste wird verworfen. Etwa alle 500 Generationen werden von den Populationen Bakterienproben entnommen und bei -80°C eingefroren. Auf diese Weise entstand ein einzigartiges Archiv von Ahnenformen – eine Datenbank, die alle wesentlichen Vorfahren der heutigen Bakterienstämme als "lebende Fossilien" konserviert. Die tiefgefrorenen Bakterien haben gegenüber stratigraphisch überlieferten Fossilien den entscheidenden Vorzug, dass sie jederzeit zum Leben erweckt und genetisch untersucht werden können.
LENSKI und Mitarbeiter ermittelten nun die Wachstumsrate in allen Bakterienpopulationen (relativ zur Ursprungspopulation). Bei konstanter Wachstumsrate dürfte sich der Stoffwechsel von E. coli kaum nennenswert verändert haben, wohingegen ein sprunghafter Anstieg der Vermehrungsrate Mutationen anzeigt, welche die Bakterien dazu befähigen, nicht nur die Glucose, sondern vor allem das im Überschuss vorhandene Citrat zu verstoffwechseln.
Eine Innovation entsteht: die Cit+- und Cit++-Mutante
Nachdem über viele Jahre kaum Veränderungen in der Wachstumsrate zu verzeichnen waren, entwickelte sich zwischen den Generationen 31.000 und 31.500 eine bemerkenswerte Neuerung: LENSKI und Mitarbeiter berichteten darüber, dass in einer der zwölf parallelen Abstammungslinien eine Population die rudimentäre Fähigkeit erwarb, Citrat aus dem Nährmedium aufzunehmen (Phänotyp Cit+ genannt) (BLOUNT et al. 2008). Bemerkbar machte sich diese Eigenschaft anhand einer drastischen Zunahme der Trübung in der als Ara-3 bezeichneten Population.
Die extrem schwache Citrat-Verwertung der Cit+-Mutante bescherte ihren Trägern jedoch praktisch keinen Fitnessvorteil (QUANDT et al. 2015). Erst ab Generation ~33.000 wurde das rudimentäre Merkmal zum stärkeren, vorteilhaften Phänotyp Cit++ weiterentwickelt, was mit einem starken Anstieg der Zelldichte in dieser LTEE-Population zusammenfiel.
Drei Etappen der Entwicklung
Es ist wichtig zu betonen, dass nicht mehrere Bakterienkulturen die Fähigkeit entwickelten, Citrat als Kohlenstoffquelle zu nutzen, sondern nur eine einzige. Das liegt daran, dass die Innovation – wie wir noch sehen werden – eine komplexe Serie spezifischer Mutationen erforderte. Zwar evolvieren die Bakterien rasch; in einer dauerhaft exponentiell wachsenden Kultur mutiert innerhalb weniger Jahre jedes Nukleotid im Genom von E. coli gleich mehrfach. Doch die Wahrscheinlichkeit, dass genau die "richtigen" Mutationen in der erforderlichen Reihenfolge auftraten, ist a priori gering.
Die ursprünglichen Analysen wurden von BLOUNT et al. (2008) und BLOUNT et al. (2012) durchgeführt. Sie entdeckten, dass die Evolution des neuen Merkmals konzeptionell oftmals in drei Entwicklungsschritte unterteilt werden kann: Ermöglichung, Verwirklichung und Verstärkung (s. Abb. 1).
Genuntersuchungen aus dem Bakterienarchiv ließen den Schluss zu, dass sich um die 20.000. Generation mindestens eine "ermöglichende" Mutation ereignet haben musste. Zwar verhalfen sie den Bakterien noch nicht zur Citrat-Verwertung. Sie schufen aber die (metabolische) Voraussetzung dafür, dass bei den Tochterbakterien eine zweite, "verwirklichende" Mutation ansetzen konnte, ohne dass dies den Bakterienstoffwechsel negativ beeinträchtigt (näheres dazu weiter unten).
Die "verwirklichende" Mutation, die sich nach rund 31.000 Generationen ereignete, bewirkte eine Neusortierung bestimmter Gene. Sie trat mehrmals unabhängig in verschiedenen Populationen auf. Allerdings hatte sie nur dort Bestand, wo sich bereits die "ermöglichenden Mutationen" ereignet hatten. Sie führte zu einem sehr schwachen Cit+-Phänotyp, der zwar in der Population über viele Generationen erhalten blieb. Aufgrund des Fehlens eines klaren Selektionsvorteils wurde sie aber nicht fixiert (QUANDT et al. 2015).
Der dritte Schritt bestand darin, durch "verstärkende" Mutationen die in Bezug auf ihre Fitness praktisch neutrale Citrat-Verwertung auf ein für das Bakterium vorteilhaftes, lebenserhaltendes Niveau anzuheben (Cit++-Phänotyp).
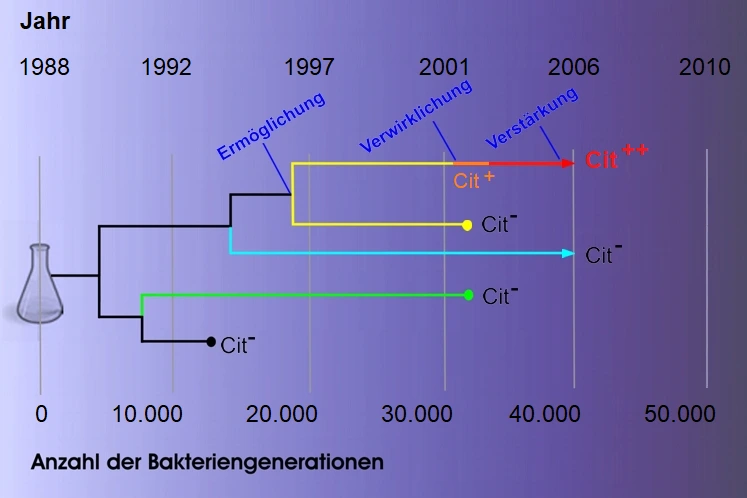
Abb. 1 Die Entstehung einer Innovation: Die ersten E. coli-Bakterien konnten Citrat noch nicht als Kohlenstoffquelle nutzen (Cit-). Dies war erst ab der 33.000 Generation möglich. Diese Evolution fand in drei Etappen statt: Im ersten Schritt schufen eine oder mehrere "ermöglichende" Mutationen die Voraussetzung für die spätere Entwicklung einer schwachen Citrat-Verwertung. Im zweiten Schritt konnte dann die "verwirklichende" Mutation ansetzen und den Cit+-Phänotyp hervorbringen. Dieser wurde schließlich zum Cit++-Phänotyp verstärkt. Modifiziert nach HENDRICKSON & RAINEY (2012).
Die ermöglichenden Mutationen
Die ermöglichenden Mutationen schufen die Voraussetzung dafür, dass eine später auftretende, rudimentäre Citrat-Verwertung (Cit+-Mutation) lange genug in der Population Bestand haben konnte, bis sie durch weitere Mutationen zur vollständigen Citrat-Verwertung (Cit++) verstärkt wurde. Bis etwa zur 20.000 Generation führte die Cit+-Mutation nämlich bei allen LTEE-Populationen zu starken Fitnesseinbußen und verschwand daher rasch wieder. Der Grund ist, dass die Aufnahme von Citrat mit dem Verlust wertvoller C4-Metaboliten einhergeht, was für die Bakterien unvorteilhaft ist.
Das Transportprotein CitT funktioniert wie eine Drehtür; es lässt Citrat in die Zelle und schleust im Gegenzug C4-Dicarboxylate, wie Succinat, Fumarat und Tartrat, aus. Wäre dieser Fitnessnachteil nicht im Vorfeld durch passende Mutationen kompensiert worden, wäre die spätere Invasion des Cit+-Phänotyps praktisch chancenlos gewesen.
Neuere Untersuchungen haben gezeigt, dass zwischen Generation 20.000 und 25.000 entscheidende Mutationen in den Schlüsselgenen gltA, iclR und arcB auftraten (QUANDT et al. 2015; LEON et al. 2018). Vor allem die gltA1- und arcB-Mutation ermöglichten es dem später auftretenden Cit+-Phänotyp, so lange in der Population zu überleben, bis der vorteilhafte Cit++-Phänotyp entstand. Wie bewirkten sie dies?
E. coli scheidet Acetat als Überschussmetabolit während des Wachstums mit Glukose aus und schaltet dann auf die Nutzung dieses Acetats um, wenn die Glukose aufgebraucht ist (WOLFE 2005). Die erwähnten Mutationen haben die Fähigkeit des LTEE-Vorfahrenstamms zur Acetat-Assimilation stark verbessert. Dadurch brachten sie die für den Wildtyp von E. coli schädliche Cit+-Mutante auf ein für die natürliche Auslese neutrales Niveau. So schuf die Optimierung des Acetatstoffwechsels beiläufig die Voraussetzungen für die Evolvierbarkeit einer rudimentären Citrat-Verwertung.
Die verwirklichende Mutation
Der Ursprung der verwirklichenden Mutation liegt in einer DNA-Region mit dem Gen citT. Dieses Gen kodiert für das Transportprotein, welches Citrat in die Zelle importiert. Ursprünglich lag citT hinter einem weiteren für die Citrat-Verwertung wichtigen Gen namens citG sowie hinter dem sogenannten rnk-Gen, das eine andere Aufgabe erfüllt.
Eine besondere Tandem-Duplikation bewirkte ein spezifisches Neuarrangement dieser Gene. Dabei geriet jeweils eine Kopie der zuvor stillen, nicht exprimierten citT- und citG-Gene unter die Kontrolle des regulatorischen Bereichs des benachbarten rnk-Gens (Abb. 2). Dieses neu entstandene rnk-citT-Modul führte zu einem neuartigen Regulationsmuster, welches die Cit-Gene auch in Gegenwart von Luftsauerstoff aktivierte und somit aerobes Wachstum auf Citrat ermöglichte (QUANDT et al. 2015).
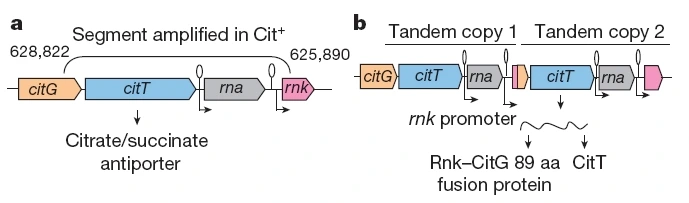
Abb. 2 a.) Räumliches und regulatorisches Arrangement des Genkomplexes (citG-citT-rna-rnk) in den ursprünglichen E. coli-Bakterien. b.) Nach einer "Tandem-Duplikation" liegt der Komplex zweimal hintereinander in der Bakterien-DNA. Der duplizierte Komplex wurde so ins Genom der Cit+-Mutante eingebaut, dass sowohl citG als auch citT unter der Kontrolle des Regulators des rnk-Gens exprimiert – das heißt auch in Gegenwart von Luftsauerstoff abgelesen – wurden. Grafik aus BLOUNT et al. (2012).
Die verwirklichende citT-Mutation des Cit+-Phänotyps wurde auch in Vorfahren dieses Stammes sowie in die anderen elf Stämme eingefügt, zeigte dort allerdings keinen Effekt. Dies belegt, dass der ab der 35.000 Generation auftretende Cit++-Phänotyp Besonderheiten aufwies, die den Cit+-Phänotyp erheblich "verstärkten".
Die verstärkenden Mutationen
Da der Cit+-Phänotyp extrem schwach ist und vermehrt essenzielle C4-Dicarboxylate ausscheidet, genügt das rnk-citT-Modul noch nicht, um dem Cit+-Phänotyp einen nennenswerten Fitnessvorteil zu bescheren. Als Bedingung für eine positive Auslese bedurfte es deshalb mindestens einer weiteren Mutation.
QUANDT et al. (2014) entdeckten eine Veränderung in der regulatorischen Region eines Gens namens dctA, ohne die die Citrat-Verwertung kaum Chancen gehabt hätte, sich durchzusetzen. Sie fanden heraus, dass die erhöhte dctA-Expression es Cit+-Zellen ermöglicht, C4-Metaboliten wie Succinat, Malat und Fumarat, die beim Import von Citrat aus der Zelle ausgeschieden werden, wieder aufzunehmen. QUANDT et al. zeigen, dass diese Mutation (in dctA) nach der ursprünglichen citT-Genmutation auftrat und dass die Kombination der beiden Mutationen für die Umwandlung des schwachen Cit+ in den starken Cit++-Phänotyp verantwortlich ist.
Nur ein einfacher Anpassungsschritt?
Erwartungsgemäß spielen Kreationisten die Tragweite des Evolutionsexperiments herunter (etwa WORT & WISSEN 2012). Sie behaupten, LENSKI habe lediglich die Regulations- oder Spezifikationsänderung eines Transportproteins nachgewiesen; keinesfalls sei in dieser Evolution etwas Neues entstanden. Dabei ignorieren sie völlig die spezifische, nicht-reduzierbare Komplexität der Mutationsanforderungen für den neuen Phänotyp:
Schritt 1: Die Mutation gltA1 bewirkte im Verbund mit weiteren Mutationen, dass sich eine spätere Citrat-Verwertung nicht nachteilig bei E. coli auswirkt.
Schritt 2: Eine spezifische Tandemduplikation brachte das rnk-citT-Modul hervor, welches die Aufnahme von Citrat aus dem Nährmedium ermöglichte (Cit+).Der Cit+-Phänotyp brachte ihren Besitzern aufgrund der verstärkten Ausscheidung von C4-Metaboliten noch keinen klaren Fitnessvorteil.
Schritt 3: Eine Mutation in der regulatorischen Region des Gens dctA verstärkte den Cit+-Phänotyp zum vorteilhaften Cit++-Phänotyp.
Kurzum: Erst nachdem mehrere Mutationen auftraten, war eine effektive Citrat-Verwertung möglich und die Selektion konnte greifen.
Die Innovation erforderte scheinbar mehrere aufeinander "abgestimmte" (konzertierte) genetische Veränderungen, die hinreichend komplex und spezifisch waren. Die Cit++-Mutante ist, um es mit HENDRICKSON & RAINEY (2012) zu formulieren, ein Schlag ins Gesicht all jener, die es für unmöglich hielten, dass die Evolution in einer Serie komplexer Ereignisse, in Zehntausenden von Generationen, neue Merkmale hervorzubringen vermag.
Zusammenfassung
Das LTEE-Experiment hat gezeigt, dass sich eine Reihe scheinbar aufeinander abgestimmter Mutationen, die in ihrer Gesamtheit ein nicht-reduzierbar komplexes Gesamtereignis konstituieren, Schritt für Schritt in einer Population akkumulieren können. Im vorliegenden Fall durchlief die betreffende LTEE-Population drei Stoffwechselepochen:
- Zunächst bewirkten verschiedene Mutationen Anpassungen im Glucose- und Acetatstoffwechsel von E. coli. Sie optimierten die Glukose-Verwertung, was eine stärkere Acetat-Akkumulation im Zellinnern zur Folge hatte.
- Danach optimierten Mutationen (wie gltA1 u.a.) die Acetat-Verwertung in Verbindung mit weiteren Verbesserungen des Glukosewachstums.
- Schließlich wurde die Citrat-Verwertung entdeckt und optimiert.
Die vorangegangenen Mutationen, die in anderen Kontexten positiv selektiert wurden, schufen unvorhersehbar und unvermittelt die Voraussetzungen für die Evolution der Citrat-Verwertung. Dabei entstand zunächst der schwache Cit+-Phänotyp, der für sich genommen noch kein selektionspositives Niveau der Citrat-Verwertung ermöglichte. Erst der starke Cit++-Phänotyp erwies sich als klar vorteilhaft.
So ist ein nichtreduzierbar komplexes System entstanden. Es entstand aber nicht durch konzertiertes Zusammenführen mehrerer unabhängiger Mutationen zum Zweck der Citrat-Verwertung. Stattdessen entstand es durch graduelle Optimierung anderer Stoffwechselprozesse, denen eine vorbereitende Brückenfunktion bei der Entstehung der Citrat-Verwertung zukam. Als die funktionelle "Brücke" nicht mehr benötigt wurde, kam es zum Funktionswechsel – und die jetzt redundant gewordenen, vorangegangenen Veränderungen im Glucose- und Acetatstoffwechsel wurden wieder eliminiert.
Dieses evolutive Schema geht mit der Beschreibung von ORR (1996) konform. ORR hat theoretisch gezeigt, wie sich durch Optimierung von Nebenwegen mit Einbettung der erworbenen Mutationen in neue Funktionszusammenhänge (Kooption) nichtreduzierbar komplexe Systeme schrittweise anbahnen können. LENSKIs Bakterienevolution bestätigt dieses theoretische Konzept eindrucksvoll.
Literatur
BLOUNT, Z.D.; BORLAND, C.Z. & LENSKI, R.E. (2008) Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli. Proceedings of the National Academy of Sciences 105, S. 7899–7906.
BLOUNT, Z.D., BARRICK, J.E., DAVIDSON, C.J. & LENSKI, R.E. (2012) Genomic analysis of a key innovation in an experimental Escherichia coli population. Nature 489, S. 513–518.
HENDRICKSON, H. & RAINEY, P.B. (2012) How the unicorn got its horn. Nature 489, S. 504–505.
LEON, D.; D’ALTON, S.; QUANDT, E.M. et al. (2018) Innovation in an E. coli evolution experiment is contingent on maintaining adaptive potential until competition subsides. PLoS Genetics 14: e1007348.
ORR, H.A. (1996) Darwin v. Intelligent Design. Boston Review 21, S. 28–31.
QUANDT, E.M.; DEATHERAGE, D.E.; ELLINGTON, A.D. et al. (2014) Recursive genomewide recombination and sequencing reveals a key refinement step in the evolution of a metabolic innovation in Escherichia coli. Proceedings of the National Academy of Sciences 111, S. 2217–2222.
QUANDT, E.M.; GOLLIHAR, J.; BLOUNT, Z.D. et al. (2015) Fine-tuning citrate synthase flux potentiates and refines metabolic innovation in the Lenski evolution experiment. eLife 2015;4: e09696.
WOLFE, A.J. (2005) The acetate switch. Microbiology and Molecular Biology Reviews 69, S. 12–50.
WORT UND WISSEN (2012) Von der Citrat-Verwertung zur Entstehung des Auges? https://www.genesisnet.info/index.php?News=187. Letzter Zugr. a. 28.02.2025.
Autor: Martin Neukamm


