
Druck-Version
Faktenwissen
Evolution auf Abwegen?
Die faszinierende Brutpflege des Froschlurchs Rheobatrachus silus
Es ist eine bekannte Strategie religiös voreingenommener Evolutionsgegner, biologische Merkmale ins Auge zu fassen, deren Entstehung bislang noch nicht im Detail geklärt worden ist. Dann wird behauptet, deren Evolution sei aus selektionstheoretischen Gründen unvorstellbar und höchst unplausibel. Dabei wird meist unterstellt, es seien zahlreiche zueinander passende Veränderungen gleichzeitig erforderlich, bis ein Selektionsvorteil gegeben sei. Aus Sicht eines "blinden" Naturprozesses wie Evolution sei dies beliebig unwahrscheinlich. Folglich müsse das betreffende Merkmal "ausgedacht" und (von Gott) intelligent geplant worden sein.
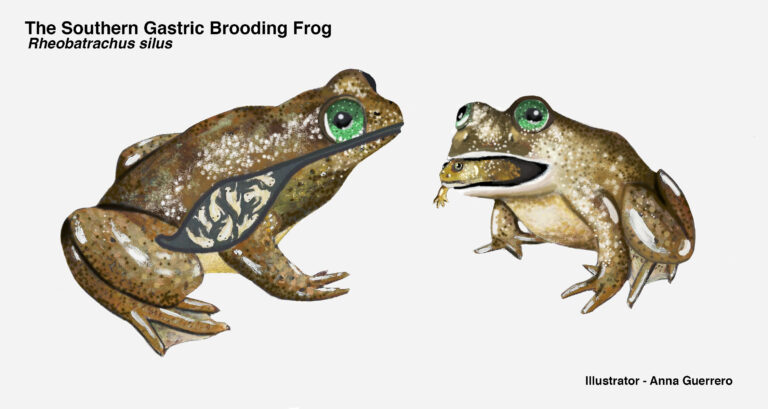
Titelbild: Zeichnung des inzwischen ausgestorbenen südlichen Magenbrüterfroschs (Rheobatrachus silus). Im Magen und im Maul der Mutter sind winzige Fröschlein zu sehen. Bildquelle: Anna Guerrero, Embryo Project Encyclopedia, CC BY-NC-SA 3.0.
Das bizarre Brutverhalten von Rheobatrachus
Als prominentes Anschauungsobjekt dient dem Kreationismus (z. B. in JUNKER & SCHERER 2013) unter anderem ein Froschlurch mit einem so ungewöhnlichen wie einzigartigen Brutverhalten: Der Magenbrüterfrosch Rheobatrachus silus ist eine von zwei extinkten, ehemals in Australien heimischen Froschlurch-Arten, deren Larven im Magen der Mutter zu jungen Fröschen heranreiften. Das Weibchen schluckte bis zu 40 besamte Eier, stellte die Verdauung ein und fastete, bis die Jungfrösche aus dem Maul geschlüpft waren. Der Magen wurde so zum Uterus für die Brut, die sich vom proteinreichen Eidotter ernährte.
Innerhalb von sechs Wochen reiften die Kaulquappen zu Jungfröschen heran; kurz vor dem Gebären erreichten sie nahezu die Hälfte des Gewichts der Mutter. Dabei komprimierte der gedehnte Magen die Lungen so stark, dass das Weibchen nur noch durch die Haut atmen konnte und seine Aktivität reduzierte. Bei der Geburt erweiterte sich die Speiseröhre des weiblichen Frosches, sodass die Jungen aus dem Maul des Muttertieres schlüpfen konnten.
Kleine Veränderungen oder großer Quantensprung?
Abgesehen davon, dass die Entstehung dieser faszinierenden Brutpflege noch nicht im Detail erforscht ist: Wo liegt hier nach Ansicht der Evolutionsgegner das Problem für die Evolutionstheorie? Angeblich darin, dass sowohl die Organfunktionen als auch das Verhalten des Froschweibchens sprunghaft große, kooperative Veränderungen erfahren mussten, damit das Ausbrüten der Jungen reibungslos funktionierte. Eine schrittweise, durch die Selektion gestützte Evolution sei, so wird uns vielfach versichert, in diesem Fall unmöglich gewesen.
Bezüglich dieser Einschätzung kommt ihnen das Statement des Zoologen und Erstbeschreibers Michael J. TYLER zupass. Dieser postuliert einen "einzelnen, großen Quantensprung" der Evolution (S. 129), weil auch er dem Anschein nach eine allmähliche Entwicklung des Magenbrutverhaltens für unmöglich hält. Neben der Unterdrückung der Magenfunktion hält er nicht weniger als 16 flankierende Veränderungen für erforderlich, um das Brutpflegeverhalten evolutionär zu etablieren (S. 132, Tab. 10.2).
In einem haben die Evolutionsgegner sicher Recht: Bestünde der einzige gangbare Entwicklungsweg in einer simultanen Ko-Anpassung verschiedenster Organe und Organsysteme ohne selektionspositive Zwischenschritte, hätte wohl nie eine Evolution dieser bizarren Brutpflege stattfinden können. Nur, und das ist das notorische Problem der Evolutionskritik, sind derlei Annahmen reine Erfindung.
Die Rolle des Hormons Prostaglandin bringt Licht ins Dunkel
Die auf den ersten Blick unerklärbare Eigenschaft des Brutverhaltens beruht auf der Tatsache, dass die befruchteten Eier offensichtlich die Magenfunktion unterdrücken. Verantwortlich dafür ist das Hormon Prostaglandin E2 (kurz: PGE2), das sowohl die Salzsäure- als auch die Pepsin-Sekretion im Magen des Muttertieres hemmt. Wird dieses Hormon von den Eiern und Froschlarven in ausreichender Menge produziert, kann das Muttertier die Eier nicht mehr verdauen, wodurch der Magen zwangsläufig zu einer Art Uterus wird.
Interessant dabei ist: Bei diesem Hormon handelt es sich um keinen evolutiven Neuerwerb der Magenbrüterfrösche, wie man vermuten könnte, sondern um ein sehr altes und sehr weit verbreitetes Gewebshormon, welches auch bei den Parietalzellen des menschlichen Magens die Salzsäureproduktion hemmt, um eine Selbstverdauung zu verhindern. Darüber hinaus kommen PGE2-Rezeptoren in der glatten Muskulatur des Uterus vor, wo das Hormon bei lebendgebärenden Wirbeltieren eine Kontraktion des Uterus bewirkt.
Bei Rheobatrachus brauchte also keine Neumutation aufgetreten sein, um die PGE2-Sekretion sicherzustellen, denn das Hormon wird auch von den Embryonen anderer Tierarten sezerniert. Falls dies nicht auch beim letzten gemeinsamen Vorfahren von Rheobatrachus der Fall gewesen sein sollte, hätte es ausgereicht, die schon existierenden Gene, die für das Hormon PGE2 kodieren, in der Larvenentwicklung "anzuschalten", um den Verdauungsvorgang im Magen des Muttertieres zu hemmen. Dies ist wohl im Wesentlichen schon die Lösung der vermeintlichen "Quantensprung-Evolution". Wenn man außerdem weiß, dass es Froschlurche gibt, die einen Teil ihrer Eier auffressen, kann das Brutverhalten der Magenbrüterfrösche als abgeleiteter Kannibalismus erklärt werden.
Anatomische Besonderheiten erleichtern die Evolution
Wie verhält es sich mit den übrigen Veränderungen, die sich angeblich zur gleichen Zeit (von einer Generation auf die andere) vollziehen mussten? Gehen wir die Punkte der Reihe nach durch, wie sie auf einer evangelikalen Website angeführt werden (ROLLER 2016):
"1. Morphologie: Möglichkeit zur Vergrößerung des Magens und zur Unterdrückung seiner Aktivität; gesteigerte Gefäßneubildung der Magenwände; Fähigkeit, durch das aufgerissene Maul die Jungen zu gebären; Fähigkeit zur maximalen Ausdehnung der Speiseröhre."
Die extreme Dehnbarkeit von Ösophagus und Magen zählt zu den anatomischen Besonderheiten der Froschlurche. Jeder, der schon einmal einen Frosch dabei beobachten konnte, wie dieser scheinbar mühelos eine ausgewachsene Maus, eine Eidechse oder einen ähnlich großen Artgenossen verspeiste, weiß dies aus Erfahrung. Dazu bemerken WESTHEIDE & RIEGER (2015, S. 313):
"Der Oesophagus ist kurz, … mit Wimperepithel ausgestattet und durch Längsfalten weit dehnbar… Der Magen ist extrem dehnbar und kann bei manchen Froschlurchen Beute aufnehmen, die der Körpermasse des Frosches selbst entspricht."
Man muss also davon ausgehen, dass die Fähigkeit zum Maulgebären allein schon durch den Bauplan der Froschlurche ermöglicht wird. Sicher war die Vergrößerung von Magen und Ösophagus ein sinnvoller Optimierungsschritt, musste aber nicht zeitgleich mit der PGE2-Sekretion aufgetreten sein. Zudem regt PGE2 die Gefäßneubildung an, indem es den vaskulären endothelialen Wachstumsfaktor induziert. Die Notwendigkeit eines entsprechenden, simultan auftretenden Ko-Evolutionsschritts entfällt damit ebenfalls.
"2. Physiologie: Unterdrückung der Magensäure-Produktion; Unterdrückung der Produktion des Verdauungsenzyms Pepsin; Erschlaffung des Darms."
Sowohl die Salzsäure- als auch die Pepsin-Sekretion werden durch PGE2 gehemmt. Dies ist der eigentliche Evolutionsschritt, der mit der PGE2-Ausschüttung initiiert wird; die Unterdrückung der Verdauungssekrete muss also nicht noch extra zu dieser "addiert" werden.
"3. Verhalten: Keine Nahrungsaufnahme während der ganzen Zeit, in der sich die Jungen im Magen befinden; passive Lebensweise".
Jeder, der schon einmal versucht hat, bei vollem Magen zusätzliche Nahrung aufzunehmen, weiß, dass dies zunehmenden Widerwillen auslöst. Man darf also getrost annehmen, dass Tiere mit überfülltem Magen zwangsläufig fasten und zu passiver Lebensweise übergehen anstatt durch zusätzliche Nahrungszufuhr zu erbrechen oder eine Perforation des Verdauungsstrakts zu riskieren.
"4. Morphologie: Vergrößerte Eidurchmesser und verringerte Fruchtbarkeit."
Auch das ist ein klassischer Fall von Optimierung: die quantitative Änderung bereits bestehender Merkmale. Warum soll dieser Anpassungsschritt von Anfang an erforderlich gewesen sein?
Kaulquappen können mehrere Wochen lang ohne Zusatznahrung überleben, bevor sie absterben. In den ersten 1-2 Wochen ernähren sie sich vom Eidotter, den Resten der gallertartigen Eiumhüllung, sowie von der in diesen lebenden Mikrofauna und -Flora. Im Anschluss daran stehen den Tieren angedaute Nahrung aus dem Magen des Muttertieres sowie verendete Larven und deren Abbauprodukte zur Verfügung. Im späteren Entwicklungsstadium, in dem den Larven Arme und Beine sprießen, dient ihnen ihr eigener Schwanz als Nahrung.
Die Annahme, die Larven hätten sich ohne vergrößerten Eidurchmesser gar nicht entwickeln können, ist somit wenig plausibel. Gewiss aber ist der vermehrte Eidotter eine bessere Starthilfe, um einer größeren Zahl von Kaulquappen das Überleben zu sichern. Eine solche Optimierung war sicher im Sinne von Dawkins "gradual ramp" möglich.
"5. Physiologie: Erzeugung von Substanzen, die die Bildung von Magensäure … unterdrücken".
Siehe Punkt 2.
"6. möglicherweise Anpassung des Atemtrakts und der »Abfall«entsorgung."
Von größter Bedeutung ist beim Frosch die Hautatmung. Für geringe Aktivitäten ist wenig Sauerstoff nötig – die Hautatmung reicht dafür völlig aus. Es gibt sogar Frösche, die benutzen fast nur die Hautatmung, die Lungenatmung ist für sie eher unwichtig.
"7. Verhalten: Verringerung der Beweglichkeit des Körpers und der Extremitäten"
Eine verringerte Beweglichkeit des Körpers ist bei Fröschen, die über die Haut atmen und daher mit weniger Sauerstoff versorgt werden als über die Lungen, systemimmanent: Frösche passen, was wenig überrascht, ihre Aktivität dem Sauerstoffgehalt in ihrem Blut an.
Im Übrigen spricht auch wenig dafür, dass dieses Brutverhalten anfangs notwendigerweise optimal funktionieren musste. Selbst wenn das Muttertier zunächst noch einige Eier verdaut haben und nur ein Teil der Jungen im Magen zu adulten Fröschen heranreifen konnte, wäre noch immer ein deutlicher Selektionsvorteil gegenüber dem Ablaichen im Wasser gegeben. Üblicherweise müssen Frösche Hunderte oder gar Tausende von Eiern produzieren, um das Überleben ihrer Art zu sichern. Bereits ein partielles Überleben im Magen des Muttertieres reduziert die notwendige Zahl an Eiern auf wenige Dutzend. Der Rest ist, wie man so schön sagt, Geschichte (sprich evolutive Optimierung).
Fazit
Evolutionsgegner neigen seit jeher dazu, durch die keineswegs originelle Behauptung, es müssten zahlreiche Änderungen gleichzeitig erfolgt sein, bevor ein Selektionsvorteil gegeben ist, die Evolution biologischer Merkmale komplizierter erscheinen zu lassen als sie ist. Allerdings konnten die Kreationisten mit dieser Strategie schon bei anderen Merkmalen nicht überzeugen (PEITZ 2013).
Auch die Art und Weise, wie sich die Kreationisten der Evolution des Magenbrüterfroschs annehmen, führt in die Irre: Alle biologischen Fakten sprechen dafür, dass der evolutive Schritt vom Magen hin zum Uterus durch die PGE2-Produktion in den Eiern und Larven erfolgte und für das Auftreten eines ersten Selektionsvorteils ausreichend war. Da alle für das Magenbrüten wesentlichen Funktionen und Strukturen der betreffenden Organe bereits bei den Vorfahren der Magenbrüterfrösche vorhanden waren, beschränkte sich die Evolution auf die quantitative Optimierung dieser Strukturen, wie etwa auf die sukzessive Vergrößerung des Eidurchmessers, die Weitung von Magen und Ösophagus sowie die Optimierung der sezernierten Menge an PGE2.
Literatur
JUNKER, R. & SCHERER, S. (2013) Evolution. Ein kritisches Lehrbuch. 7. Aufl., Weyel, Gießen.
PEITZ, H.-H. (2013) Kreationistische Rückzugsgefechte.
ROLLER, T. (2016) Wie ein Frosch die Evolutionstheorie ins Wanken bringt.
TYLER, M. J. (1983) The gastric brooding frog. Croom Helm, London & Canberra.
WESTHEIDE, W. & RIEGER, G. (2015) Spezielle Zoologie. Teil 2: Wirbel- oder Schädeltiere. 3. Aufl., Springer, Berlin.
Nachtrag vom 16.08.2016: Anmerkungen zu einer Kritik von T. ROLLER
Vor ein paar Wochen hat Timo ROLLER von der Studiengemeinschaft WORT UND WISSEN auf meinen Beitrag geantwortet. Im Folgenden möchte ich einige Überlegungen zu seiner Kritik anstellen.
1. Autoritätsargumente sind keine Begründungen
ROLLER erachtet es zunächst als nötig, seine Leser darauf hinzuweisen, dass sich meine Ausführungen de facto nicht gegen eine Erfindung der Evolutionsgegner, sondern gegen ein Szenario des Evolutionsbiologen M. TYLER richten, der für das Magenbrüten nicht weniger als 16 Anpassungen für erforderlich hält. Er versteigt sich dabei zu der Behauptung, ich hätte TYLERs Szenario fälschlicherweise als Produkt seiner evolutionskritischen Abhandlung ausgewiesen. Wie er darauf kommt bleibt unklar – in meiner Kritik wird das 16-Fach-Ereignis jedenfalls unmissverständlich Michael TYLER zugeschrieben und lediglich erwähnt, dass dieses Szenario "auf einer evangelikalen Website" angeführt wird:
"Eine schrittweise, durch die Selektion gestützte Evolution sei, so wird uns vielfach versichert, auch in diesem Fall unmöglich. Bezüglich dieser Einschätzung kommt ihnen [den Kreationisten] das Statement des Zoologen und Erstbeschreibers Michael J. TYLER zupass. Dieser [sic!] postuliert einen 'einzelnen, großen Quantensprung' der Evolution (S. 129), weil auch er [sic!] dem Anschein nach eine allmähliche Entwicklung des Magenbrutverhaltens für unmöglich hält." (Hervorhebung nicht im Original.)
Zudem gewinnt man den Eindruck, ROLLER wolle mit dem Verweis auf TYLERs Autorität dessen ungewöhnliche Auffassung plausibler erscheinen lassen (Autoritätsargument). Doch ist die Notwendigkeit eines unwahrscheinlichen Simultanszenarios einleuchtender, wenn es von einem einzelnen Forscher postuliert wird, der sich mit der Evolutionsfrage sonst nicht näher beschäftigt? ROLLERs Feststellung, der Experte werde schon wissen, warum das so sei, ist jedenfalls kein Argument, sondern eine so genannte Killerphrase.
2. TYLER ist kein Evolutionsgegner
Auch der Versuch, TYLERs Argumentation für die Evolutionskritik zu vereinnahmen, führt in die Irre. TYLER denkt gar nicht daran, biologische Merkmale ins Auge zu fassen, deren Entstehung bislang noch nicht im Detail geklärt worden ist, um zu folgern, ihre Evolution sei nicht plausibel. Dies ist eine (logisch unhaltbare) Strategie, der sich exklusiv die religiös motivierten Evolutionsgegner verschrieben haben.
Wie immer man zu TYLERs postuliertem "Quantensprung der Evolution" steht – schon der Wortsinn zeigt es auf: Im Gegensatz zu den Kreationisten setzt TYLER eine Evolution voraus! Darüber hinaus bleibt völlig offen, was er konkret vertritt. Ob TYLER den Vollzug der Anpassungsschritte auf unzählige aufeinander "abgestimmte" Einzelmutationen zurückführt, ist keineswegs ausgemacht. Er könnte genauso gut auch eine oder einige wenige "Systemmutationen" GOLDSCHMIDT'schen Typs im Kopf haben oder verschiedene Voranpassungen und eine hohe ontogenetische Plastizität voraussetzen, die eine Evolution erleichtern.
Solange nicht klar ist, von welcher Seite sich der Biologe dem postulierten Quantensprung der Evolution nähert und welche Plausibilitätsabschätzungen er vornimmt, ist für die Evolutionskritik nichts gewonnen – sie kann lediglich die Unplausibilität eines 16-fach-Simultanereignisses feststellen. ROLLER hätte sich und seinen Lesern das unfruchtbare Vorhutgefecht also ersparen und gleich in medias res übergehen sollen.
Ist seine Sachkritik überzeugender als seine rhetorischen Stilmittel?
3. Ein Zusammenspiel von Merkmalen impliziert nicht ihre simultane Entstehung
Zunächst bemängelt ROLLER:
"NEUKAMM behandelt die meisten der Veränderungen unabhängig voneinander und beachtet nicht das ausgeklügelte Zusammenspiel dieser Eigenschaften, das ja gerade TYLER in seinen Untersuchungen feststellt. Diese isolierte Betrachtung ist biologisch unsachgemäß und unrealistisch."
Offensichtlich hat ROLLER missverstanden, worum es geht, zudem erhebt er eine Falschbehauptung: Meine Argumentation lässt keineswegs das "Zusammenspiel dieser Eigenschaften" außer Acht. Es ging um etwas anderes, nämlich um die Widerlegung der These, dass all diese Eigenschaften gleichzeitig entstehen mussten! Der Bauplan der Frösche zeichnet sich per se durch einen Großteil jener Merkmale aus, die gemäß TYLER erst de novo evolvieren mussten. Wie der Zoologe weiß, sind sowohl die Dehnbarkeit von Magen und Oesophagus als auch die Kompressibilität der Lunge, die Fähigkeit zur Hautatmung und vieles andere günstige Voranpassungen, die eine Evolution dieses außerordentlichen Brutverhaltens wahrscheinlich überhaupt erst ermöglicht haben.
Fern biologischen Tatsachenwissens ist auch ROLLERs Behauptung, das Muttertier müsse in irgendeiner Weise darauf "vorbereitet" gewesen sein, "von einer Generation auf die nächste anders als zuvor sechs Wochen lang ohne Nahrungszufuhr auskommen, bis die Frösche ausreichend entwickelt waren…". Es ist doch schon lange bekannt, dass wechselwarme Tiere wie Schlangen, Eidechsen und Amphibien nicht nur mehrere Wochen, sondern monatelang ohne jede Nahrung auskommen, ohne zu sterben. Falls es seitens Rheobatrachus überhaupt einer "Vorbereitung" auf diesen Ausnahmezustand bedurft haben sollte, dann ist diese in seinem genetischen Erbe zu finden, nicht in einer dubiosen Vorausplanung biologischer Merkmale durch ein so unbekanntes wie unerforschliches Wesen.
4. Wesentliche 'Passungen' existieren bei Froschlurchen bereits 'von Haus aus'
Weiterhin moniert mein Kritiker, NEUKAMM vergleiche Rheobatrachus
"mehrmals mit irgendwelchen Fröschen (z. B. die Dehnbarkeit von Speiseröhre und Magen, Überlebensfähigkeit von Kaulquappen). Sachgemäß wäre ein Vergleich mit solchen Fröschen, die realistischerweise als Modelle für die nicht magenbrütenden Vorläufer von Rheobatrachus gelten können."
Das ist nun wirklich grober Unfug, denn die besonderen Merkmale sind kennzeichnend für den Bauplan der (sprich: aller) Frösche. Da die (unbekannten) Vorfahren von Rheobatrachus mutmaßlich ebenfalls Frösche waren, darf man voraussetzen, dass auch sie die entsprechenden Merkmale besaßen – es sei denn, es gibt Belege, die dagegensprechen.
Kurzum: Da wesentliche Voraussetzungen und Strukturen bereits im System etabliert sind, konzentriert sich die Evolution im Wesentlichen auf der Optimierung dieser Strukturen – es kam hauptsächlich darauf an, das Hormon PGE2 in den Froscheiern zu sezernieren, wodurch die Verdauung gestoppt wird. Im Gegensatz dazu behauptet ROLLER, die Unterdrückung der Magensäureproduktion "mache noch lange kein Magenbrüten, denn der Frosch muss gleichzeitig sein Ernährungsverhalten radikal ändern." Aber wo ist der Beleg dafür? Dass ein Frosch, der den Bauch voller unverdauter Eier hat, nicht weiter Nahrung zu sich nimmt, sollte einleuchten.
ROLLER meint des Weiteren, der Magenbrüterfrosch müsste der Theorie zufolge "eines Tages zufällig ausnahmsweise Eier fressen…" Warum ausnahmsweise? Wie erwähnt gibt es Froschlurche, die einen Teil ihrer Eier auffressen, sodass das Brutverhalten der Magenbrüterfrösche als abgeleiteter Kannibalismus erklärt werden kann (was übrigens auch TYLER vorschlägt).
5. Froschlurche sind hart im Nehmen
In ROLLERs Replik scheint die Ernährungsfrage eine zentrale Stellung einzunehmen. Es sei beispielsweise fraglich, ob die im Mutterleib heranreifenden Jungfrösche überhaupt "eine Überlebenschance gehabt hätten, wenn ihre Entwicklung im Magen mit wenig eigenem »Startkapital« (sprich: mit kleinen Eiern) begonnen hätte". Mit einem Durchmesser von 4,6 bis 5,1 mm seien die Eier von Rheobatrachus deutlich größer als die Eier vergleichbarer Froscharten. "NEUKAMM", so mein Kontrahent, müsse sich "mit weiteren unrealistischen Szenarien behelfen:
"»Selbst wenn das Muttertier zunächst noch einige Eier verdaut haben und nur ein Teil der Jungen im Magen zu adulten Fröschen heranreifen konnte, wäre noch immer ein deutlicher Selektionsvorteil gegenüber dem Ablaichen im Wasser gegeben.« – Wie soll ein Teil der Eier im selben Magen verdaut worden sein, andere Eier sich dagegen vollständig entwickelt haben? Außerdem würde ein teilweises Verdauen dann doch den Appetit erneut anregen und der Hormonspiegel von PGE2 würde fallen; NEUKAMMs Szenario ist daher in sich widersprüchlich."
Was den Größenvergleich hinsichtlich der Froscheier anbelangt, so orientiert sich ROLLER bei nicht maulbrütenden Fröschen erkennbar am unteren Ende der Skala, spricht aber von einem "Maximaldurchmesser von 1,5 mm". Nimmt man den Grasfrosch als Beispiel, sind die Eier (ohne umgebende Gallerte) zwischen 1,7 und 2,8 Millimeter groß. Auch die Eier anderer Nicht-Maulbrüter sind selten kleiner als 1,5 mm. Dies ist noch immer weniger als bei Rheobatrachus, aber die implizite Annahme, dass die kleinen Kaulquappen ohne zusätzliches "Startkapital" binnen weniger Wochen abstürben, ist abenteuerlich.
Erstens steht den Larven die Gallerte zur Verfügung, zweitens angedaute Zusatznahrung aus dem Muttermagen. Drittens überleben per se nicht alle Kaulquappen (die toten Exemplare werden ebenfalls gefressen). Und viertens stoßen die Larven nach wenigen Wochen ihren eigenen Schwanz ab, der als weitere Nahrungsquelle dient. Weshalb es sich dabei um ein unrealistisches Szenario handeln soll, wird überhaupt nicht klar; alle hier gemachten Annahmen sind durch empirische Beobachtungen untermauert.

Ausgewachsene Frösche und Froschlarven (hier: eines Laubfroschs) sind hart im Nehmen. Die erwachsenen Tiere kommen monatelang ohne Nahrung zurecht; die Larven können sich von Pflanzenresten, angedauter Nahrung und später von ihrem eigenen Schwanz ernähren. Trotzdem entwickeln sich aus bis zu 40 Eiern, die das Muttertier von Rheobatrachus frisst, nur etwa 25 Jungfrösche. Offenbar verendet ein Teil der Larven im Muttermagen, der den übrigen Kaulquappen als Zusatznahrung dienen kann. Die meisten Einwände, die ROLLER gegen eine Evolution der Magenbrüter ins Feld führt, sind unbegründet. Bildquelle: M. Linnenbach, Laubfrosch Larve, CC BY-SA 3.0.
{kind=link}
6. Die Unterdrückung des Verdauungsvorgangs muss nicht optimal sein
Auch der angebliche Widerspruch in meinem Szenario ist konstruiert. Auf ROLLERs rhetorische Frage, wie ein Teil der Eier im selben Magen verdaut worden sein solle, während sich andere Eier vollständig entwickelt haben, gibt es eine einfache Antwort: Bei der Aufnahme der Froscheier landen diese in einem noch voll funktionsfähigen Magen. Dass es eine Zeit lang dauert, bis genügend PGE2 abgesondert und der Protektions-Mechanismus in Gang gesetzt wurde, ist einleuchtend. Dadurch wird ein Teil der Eier stärker durch Magensäure und Verdauungsenzyme angegriffen (bzw. angedaut) als die übrigen.
In der Tat reifen nicht alle Larven zu adulten Fröschen heran: Obwohl das Muttertier bis zu 40 Eier aufnimmt, entwickeln sich nur etwa 20 bis 25 bis zur Geschlechtsreife. Optimalität ist etwas anderes. Verhindert werden könnte dieses Larvensterben u. a., wenn das Muttertier seine Verdauung bereits vor der Eiaufnahme stoppte und insgesamt weniger Eier aufnähme. Da aber die Evolution nicht vorausplant, sind solche suboptimalen Lösungen zu erwarten.
In diesem Stil geht es weiter: Das Muttertier hätte "... gleich so viele Eier auf einmal" fressen müssen,
"... dass es keinen weiteren Appetit mehr hatte. Die verschluckten Eier müssten glücklicherweise genügend PGE2 ausgesondert haben, damit die Magensäureproduktion vollständig gestoppt wurde. Weiter muss PGE2 von Anfang an ziemlich exakt die passenden sonstigen Auswirkungen gehabt haben, was unrealistisch ist."
Auch diese Einwände sind nicht mehr als eine Aneinanderreihung von Killerphrasen: Zum einen kann ROLLER nicht wissen, ob oder wie viel "Glück" es brauchte, damit die Eier "genügend" PGE2 sezernierten. Bei Hormonen reicht es meist aus, wenn die empfangende Zielzelle einige wenige Moleküle auffängt, um die Wirkung zu entfalten. Bereits kleinste Mengen PGE2 dürften genügen, um den Verdauungsprozess zu stoppen – und eine anfangs "versehentlich" deutlich höhere Dosierung endet nicht zwangsläufig letal. Mit "Glück" im Sinne eines unwahrscheinlichen Lotteriegewinns hat das Ganze also herzlich wenig zu tun.
Zum anderen bleibt nebulös, was ROLLER unter "ziemlich exakt die passenden sonstigen Auswirkungen" des PGE2 verstanden haben will. Wie erwähnt bewirkt das Hormons PGE2 eine ganze Kaskade physiologischer Veränderungen, die das Magenbrüten begünstigen. Das ist keine "unrealistische" Behauptung, sondern eine empirisch belegte Tatsache: Durch PGE2 werden sowohl die Salzsäure- als auch die Pepsin-Sekretion gehemmt. Zudem regt PGE2 die Gefäßneubildung an, indem es den vaskulären endothelialen Wachstumsfaktor induziert. Dies sind Auswirkungen, die sich auch an anderen Fröschen beobachten lassen.
Nebenbei bemerkt: Genau hier zeigt sich das Prinzip der "erleichterten Variation" – die enorme Plastizität des Merkmals-Tinkerings, die beim Neuverschalten bereits existierender Kernprozesse, beim Einbetten einzelner Bausteine aus einem vielfältigen Repertoire des entwicklungsgenetischen "Werkzeugkastens" in andere zelluläre Prozesse und Entwicklungskaskaden zum Tragen kommt: Ein Gewebshormon, das vielgestaltige Aufgaben übernehmen kann, wird unversehens in einem anderen Zusammenhang exprimiert und schafft dadurch die Voraussetzungen für die Entstehung eines völlig neuen Bauplans.
7. Eine graduell höhere Überlebensquote der Jungtiere im Magen gegenüber ihrer Entwicklung im freien Gewässer ist ausreichend
ROLLER begeht indes den immer gleichen Fehlschluss der Evolutionskritik, das Endergebnis einer langen Serie von Optimierungsschritten mit einem "Minimalzustand" eines funktionierenden Anfangszustandes gleichzusetzen. So weist er auf verschiedene anatomische Besonderheiten von Rheobatrachus hin, wie auf das asymmetrisch gelegene Ovar, die besonderen Harnkanäle und den Brutsack, den das Muttertier während der Fortpflanzungszeit aus einem Teil ihres Magens formt, usw. Dies sind Merkmale, die Rheobatrachus zweifelsohne für das Magenbrüten optimiert hat. ROLLERs implizite Mutmaßung jedoch, dass ohne all diese Besonderheiten das Magenbrüten von vorn herein zum Scheitern verurteilt wäre, müsste er beweisen! Sie erscheint genauso unbegründet wie seine übrigen Einwände.
Optimalität ist beileibe nicht gleichzusetzen mit (funktionaler) Nichtreduzierbarkeit und Suboptimalität impliziert nicht per se, dass der gesamte Prozess scheitert, andernfalls wären nicht einmal Anpassungsprozesse möglich, die Kreationisten belieben, "Mikroevolution" zu nennen. Dass der hochoptimierte "Komplex" von Anfang an für das Magenbrüten vorhanden gewesen sein musste, wird nur behauptet – einen Nachweis dafür gibt es nicht.
8. Das Fehlen einer Erklärung ist kein Argument für 'Design'
Problematisch ist vor allem, dass ROLLER aufgrund des (vermeintlichen oder tatsächlichen) Fehlens einer stringenten evolutionären Erklärung der so genannten "Schöpfung" einen Blankoschein ausstellt. Obwohl der postulierte Schöpfer und dessen Wirkmechanismen so unbekannt wie unerforschlich sind, sodass das Schlagwort der "Schöpfung" nicht das Geringste zur Erklärung dessen beiträgt, was erklärt werden soll, folgert ROLLER,
"… dass der Magenbrüterfrosch nur als ein einzigartiges Geschöpf Gottes zu erklären ist und auf wunderbare Weise zeigt, was Gott gemeint hat".
Kann Evolutionskritik allen Ernstes diese metaphysische Aussage rechtfertigen? Nein; ROLLERs Schlussfolgerung ist nichts weiter als das altbekannte argumentum ad ignorantiam – ein Fehlschluss, nach dem fehlendes Wissen dazu benutzt wird, um eine wissenschaftliche Theorie infrage zu stellen, und zugleich eine andere (konkurrierende) Theorie plausibler erscheinen zu lassen als sie ist.
Würden Wissenschaftler so argumentieren, gäbe es nichts mehr zu erforschen, denn dann stünde von vorn herein fest, dass Gott für alle Entstehungsphänomene, über die wir derzeit noch nichts oder zu wenig wissen, direkt verantwortlich sei. Damit begeht ROLLER die intellektuelle Todsünde des ignotum per ignotius: Etwas Unbekanntes und nach ROLLER "Hochspekulatives" wird durch etwas noch Unbekannteres "erklärt", dessen Existenz überhaupt nicht mehr empirisch begründet, geschweige denn überprüft werden kann. Damit zeigt sich, dass sein Erklärungsansatz genau das ist, was der Kreationismus immer bestritten hat: Er ist mit der rational-wissenschaftlichen Erklärungsstrategie unverträglich.
Autor: Martin Neukamm


