![]()
PDF-Version
Buchbesprechung
Alan Feduccia (2020): 'Romancing the Birds and Dinosaurs'
Brown Walker Press, 336 Seiten, ISBN 1599426064, Preis: 31,60 €
Darren Naish1)

Seit vier Jahrzehnten behauptet Dr. Alan FEDUCCIA von der University of North Carolina, dass sich alle Welt über die Dinosaurier irre. FEDUCCIAs Hauptthese lautet, dass Vögel sich nicht aus theropoden Dinosauriern entwickelt haben könnten, sondern ihren Ursprung in kletternden, vierfüßigen Nicht-Dinosaurier-Reptilien der Trias hätten. Diese Meinung bringt er in Fachartikeln und Meinungsbeiträgen zum Ausdruck, vor allem aber in seinen Büchern The Origin and Evolution of Birds (FEDUCCIA 1996) und Riddle of the Feathered Dragons (FEDUCCIA 2012). Er vertritt diese Ansicht, weil er die Theropoden als zu spezialisiert für ein terrestrisches, aufs Laufen ausgerichtetes Leben ansieht und weil er meint, dass Vögel, Vogelflug und Federn sich nur in bewaldeter Umgebung entwickelt haben könnten. In diesem Szenario entwickelte sich der aktive Schlagflug über das Stadium eines Gleitflugs von den Bäumen herab.
Zu FEDUCCIAs Leidwesen ist das "Vögel-sind-Theropoden"-Paradigma in der modernen Dinosaurierforschung fest verankert. Nicht nur, dass die Entstehung der Vögel als ein bedeutendes Ereignis in der Geschichte der Dinosaurier wahrgenommen wird; darüber hinaus wurden Elemente der Anatomie, des Verhaltens und der Biologie der Vögel in Dinosauriergruppen aller Art identifiziert. Anders ausgedrückt: Die Ansicht, dass Nichtvogel-Dinosaurier eng mit den Vögeln verwandt sind, ist Teil des Mainstream-Denkens.
Das Buch Romancing the Birds and Dinosaurs: Streifzüge durch die postmoderne Paläontologie (RTBAD) ist weder eine Anleitung für Paläo-Fans noch enthält es eine Hommage oder einen Verweis auf den Film Romancing the Stone von 1984. Vielmehr besteht es aus 23 Essays zum Stand der Dinosaurierforschung, so wie FEDUCCIA sie sieht. Der Titel ist von FEDUCCIAs Idee inspiriert, dass er auf John OSTROMs Publikation Romancing the Dinosaurs aus dem Jahr 1987 antwortet. Die Titelwahl ist etwas seltsam, da OSTROMs Artikel weder als bekannt noch als besonders einflussreich gelten kann. Zudem sind Diskussionen zur Herkunft der Vögel zwar Gegenstand von FEDUCCIAs Buch. Aber das ist nicht sein Hauptthema, denn ein Drittel des Inhalts (Kapitel 1 bis 7) befasst sich mit der Kritik an der Arbeit über die Physiologie der Dinosaurier, darüber, was Dinosaurier sind, und über den Hype, den die Dinosaurier seit ihrer Popularisierung in den 1960er bis 1990er Jahre zusehends erleben. Die Kapitel 8 bis 11 sind hauptsächlich strittigen Fragen der Vogelevolution gewidmet, wie etwa der Paläognathen-Biogeographie, dem Pädomorphismus und der Homologie der Phalangen.
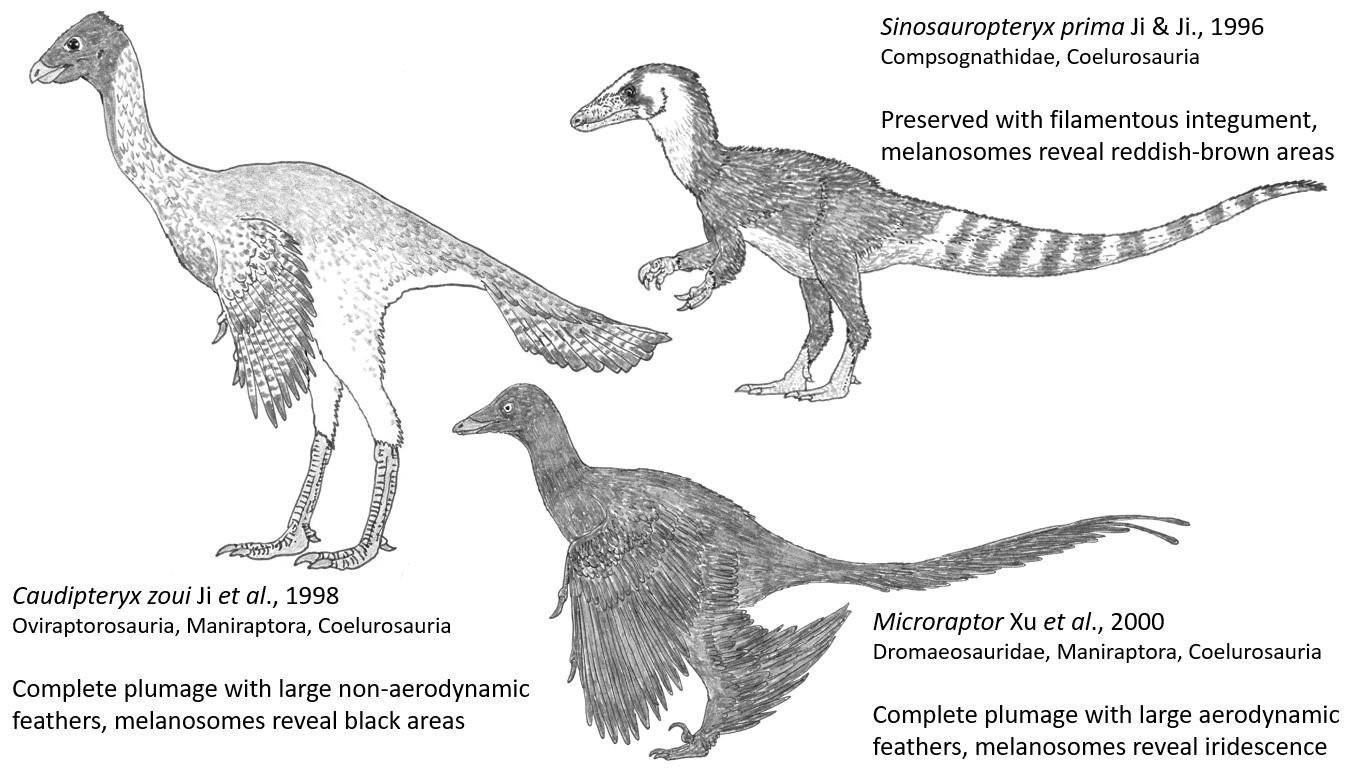
Einige wichtige Theropoden-Taxa, die in RTBAD eingehend diskutiert werden. Sie alle zählen zur Theropoden-Gruppe der Coelurosaurier. Caudipteryx und Microraptor gehören zur Coelurosaurier-Untergruppe der Maniraptoren. Bild: Darren NAISH.
Schließlich werden in den Kapiteln 12 bis 21 die gefiederten Theropoden des Jura und der Kreidezeit neu interpretiert. Theropoden mit haarähnlichem Körperbewuchs, wie etwa der kleine Sinosauropteryx, hätten, so FEDUCCIA, keine keratinösen (federähnlichen) Hautanhängsel besessen, sondern seien in Wahrheit mit Kollagenfasern bedeckt gewesen. Vogelähnliche Nichtvogel-Theropoden (Maniraptoren; MN & AB), wie etwa der truthahnähnliche Caudipteryx und der vierflügelige Microraptor, seien, so behauptet FEDUCCIA, in Wahrheit verkannte Vögel oder (wie im Falle der kleinen Scansoriopterygiden) überhaupt keine Theropoden. Die beiden letzten Kapitel enthalten Gedanken zum Aussterben am Ende der Kreidezeit, eine Zusammenfassung und so etwas wie einen "Ruf zu den Waffen".
Worum genau geht es in diesem Buch?
RTBAD lässt sich als Versuch beschreiben zu zeigen, dass die von Fachleuten vertretene Sicht auf die Dinosaurier chaotisch, oberflächlich und unreflektiert sei. Sie werde vom Streben nach Popularität und dem Festhalten an einem "kultähnlichen" Kladismus2) bestimmt. Nun feiern die meisten modernen Bücher über fossile Tiere das aktuelle, goldene Zeitalter paläontologischer Entdeckungen. Im Gegensatz dazu ist RTBAD von Negativität durchdrungen und vertritt die Ansicht, dass sich die Paläontologen größtenteils in allem irren.
Besagte Paläontologen seien Teil einer Verschwörung, in der alle wesentlichen Arbeiten über vogelähnliche Dinosaurier und dinosaurierähnliche Vögel durch eine krasse Form der Bestätigungsverzerrung3) angetrieben würden: FEDUCCIA unterstellt den Paläontologen, eine "theoriebeladene", "monolithische" Wissenschaft zu praktizieren und stellt wiederholt fest, dass Schlüsse gezogen würden, um "Kladogramme passend zu machen", nur damit sie zu den vorgegebenen phylogenetischen Erzählungen passen.

Eines muss man Alan FEDUCCIA lassen: Seine Bücher enthalten einige wunderschöne Zeichnungen; ich denke dabei insbesondere an die Darstellungen von John P. O'NEILL. Das Buchcover zeigt dieses hervorragende Bild eines fruchtfressenden Sapeornis. Rechts daneben sieht man einen terrestrischen Dromaeosauriden auf Nahrungssuche. Bild: © Zhiheng LI, Institut für Wirbeltierpaläontologie und Paläoanthropologie. Abdruck mit freundlicher Genehmigung.
FEDUCCIA übersieht die Ironie, dass seine eigene Forschung tatsächlich von einer noch gezielteren Form der Bestätigungsverzerrung angetrieben wird als die, die er fälschlicherweise bei anderen identifiziert. In seinem Bemühen zu zeigen, dass die Vogelhand nicht der von Theropoden homolog sei, dass es sich bei den Hautanhängen der Theropoden um Kollagenfasern handele und dass Caudipteryx und andere gefiederte Dinosaurier zu den Vögeln gehören, ist FEDUCCIAs Forschungsprogramm schon immer einseitig darauf ausgerichtet gewesen, sein "Vögel-sind-keine-Dinosaurier"-Paradigma zu bestätigen. Diesbezüglich war er jedoch erfolglos, wie er selber einräumt, und beklagt, dass sein vorheriges Buch "bei Paläontologen nie populär wurde" (S. 311).
Da die Wissenschaft Skeptizismus, Fehlerkorrektur und unkonventionelles Denken begrüßt, lässt sich argumentieren, dass FEDUCCIAs Rolle wertvoll, vielleicht sogar notwendig sei. Ohnehin vertritt FEDUCCIA im gesamten Buch die Meinung, dass er und eine ausgewählte Gruppe von Kollegen die wahre Wissenschaft repräsentiere. Sie seien diejenigen mit der richtigen Methodik, jene, die einen aufrichtigen Skeptizismus praktizierten und die unabhängig von populären zeitgenössischen Trends arbeiteten. Sie allein könnten den Leser sicher durch das launische, oberflächliche, chaotische Gewirr an Behauptungen über Dinosaurier und archaische Vögel führen, die von den heutigen wissenschaftlich naiven, machthungrigen, jugendlichen Populisten aufgestellt würden. Einige dieser Leute, so lesen wir, "haben Twitter-Accounts mit großen Followern [sic!], die sich mit allem befassen, von paläontologischen Entdeckungen bis hin zu Sport und Politik!" (S. 21). Nimm das, Steve BRUSATTE!4)
Das Vorwort und die ersten Kapitel von RTBAD sind eine Art Mahnruf an die moderne Paläontologie (Kapitel 1: "Brennende Dunkelheit: die neue theoriebeladene Fossilienforschung"; Kapitel 2: "Der Weg zur paläontologischen Postmoderne"; Kapitel 3: "Mach es neu! Die Dinosaurier-Renaissance"). Ich selbst gehöre zu den schrecklichen Menschen, die FEDUCCIA hier im Sinn hat und werde auch entsprechend gescholten. Besonders amüsiert mich, dass FEDUCCIA offenbar nicht in der Lage war, einen Artikel, den ich am 1. April veröffentlicht habe (Mass survival of multitudinous dinosaur lineages across the K-Pg Boundary), richtig zu verstehen.
Alan FEDUCCIA, ich klage an
So sehr ich auch versuche, den Autor als gelehrte, dem Leser wohlgesonnene Koryphäe zu betrachten, Tatsache ist, dass RTBAD geradezu karikaturhaft mit Vorurteilen und schmutzigen Tricks überladen ist: Whataboutismus (siehe unten), intellektuelle Unredlichkeit, Wiederholung falscher Behauptungen, Strohmann-Argumente und Milchmädchenrechnungen tauchen überall auf, wo der Autor versucht, gegen unliebsame Positionen zu argumentieren. Tatsächlich spiegelt sich in seinen vehementesten Argumenten nichts weiter als subjektiver (Nicht-) Glaube wider.
Der Whataboutismus ist übrigens eine Argumentationstechnik, bei der Sachverhalte, die mit dem eigentlichen Thema nichts zu tun haben, benutzt werden, um zu versuchen, die Position des Gegners zu schwächen. Ja, der zusammengesetzte "Archaeoraptor" und der falsch identifizierte Oculudentavis (S. xii-xv) stellen in der Tat krasse Irrtümer dar, und es stimmt auch, dass der taxonomische Status einiger Theropoden-Fossilien umgedeutet wurde, manchmal sogar mehrmals (S. 5-6). FEDUCCIA will damit zeigen, dass die verantwortlichen Wissenschaftler versuchen, sich als "neue Autoritätspersonen" (S. xii) aufzuspielen und dass eine Art kladistischer Fundamentalismus das Ruder übernommen habe. Ich hingegen sehe nur Wissenschaftler, die versuchen, Wissenschaft zu betreiben und dabei manchmal Fehler machen. Es ist nicht fair zu sagen, dass sich FEDUCCIA wie ein Kreationist aufführe. Aber es ist schwierig diesen Vergleich zu vermeiden, wenn man bedenkt, dass der wiederholte Verweis auf "Archaeoraptor" dem ewigen Trick der Kreationisten, die Piltdown-Fälschung aufzuwärmen, furchtbar ähnelt.
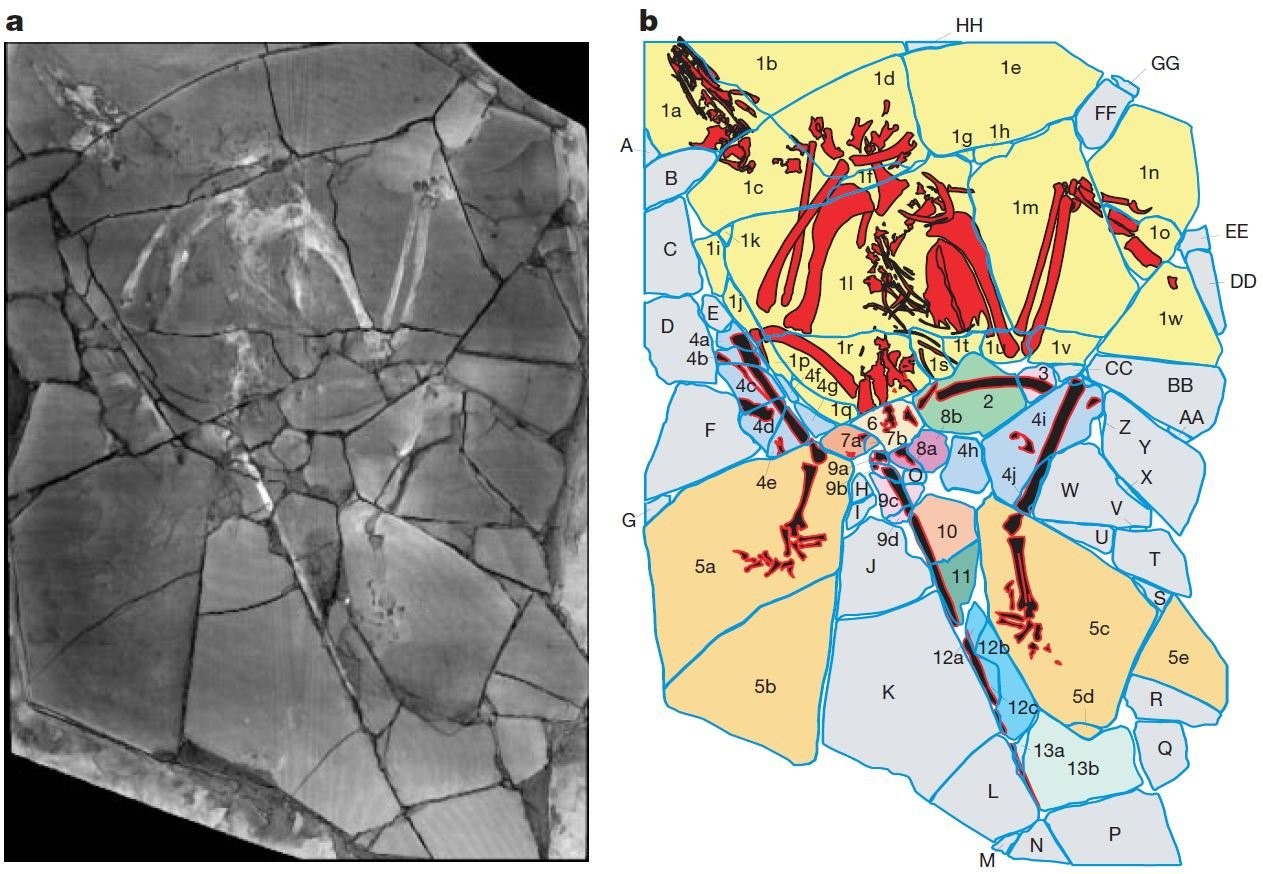
In populären Medien wie dem National-Geographic-Magazin wurde der "Archaeoraptor" als bedeutsames Mosaiksteinchen im Verständnis der Dinosaurier- und Vogelevolution gepriesen. ROWE et al. (2001) konnten jedoch nachweisen, dass es sich um eine aus verschiedenen Tierarten zusammengefügte Chimäre handelte, um ein gefälschtes Fossil also. [MN & AB: Es gibt chinesische Fossilienhändler, die Fossilien manipulieren, um höhere Verkaufserlöse zu erzielen.] Welche Auswirkungen hatte dieser peinliche Fehler auf Studien über archaische Vögel und ihrer Dinosaurier-Verwandten im Allgemeinen? Um es kurz zu machen, gar keine. Die einzigen, die diese falsche Fährte legen, sind die Kreationisten. Und natürlich Alan FEDUCCIA. Bild: ROWE et al. (2001).
Besonders aufschlussreich sind die Wiederholungen, mit denen FEDUCCIA arbeitet. Wenn man erreichen möchte, dass jemand eine These akzeptiert, die (bestenfalls) umstritten oder (schlimmstenfalls) falsch ist, muss man sie nur ständig wiederholen und hoffen, dass sie schlussendlich akzeptiert wird.
FEDUCCIA möchte seinen Lesern unbedingt weismachen, dass das angeblich "gefiederte" triassische Longisquama über Flugeigenschaften verfügte und für die Herkunft der Vögel von Bedeutung sei. Bei jeder Erwähnung des Tiers wird die Vorstellung kolportiert, dass es sich um einen "Gleitflieger" gehandelt habe. So heißt es im Brustton der Überzeugung, "... der triassische Gleitflieger Longisquama, ein kleiner baumbewohnender Archosaurier, der Fallschirmsprünge absolvierte..." (S. 87). Nun ist die Hypothese, dass Longisquama Federn zum Gleiten besessen haben könnte, zwar interessant, sie ist aber alles andere als wohlfundiert.
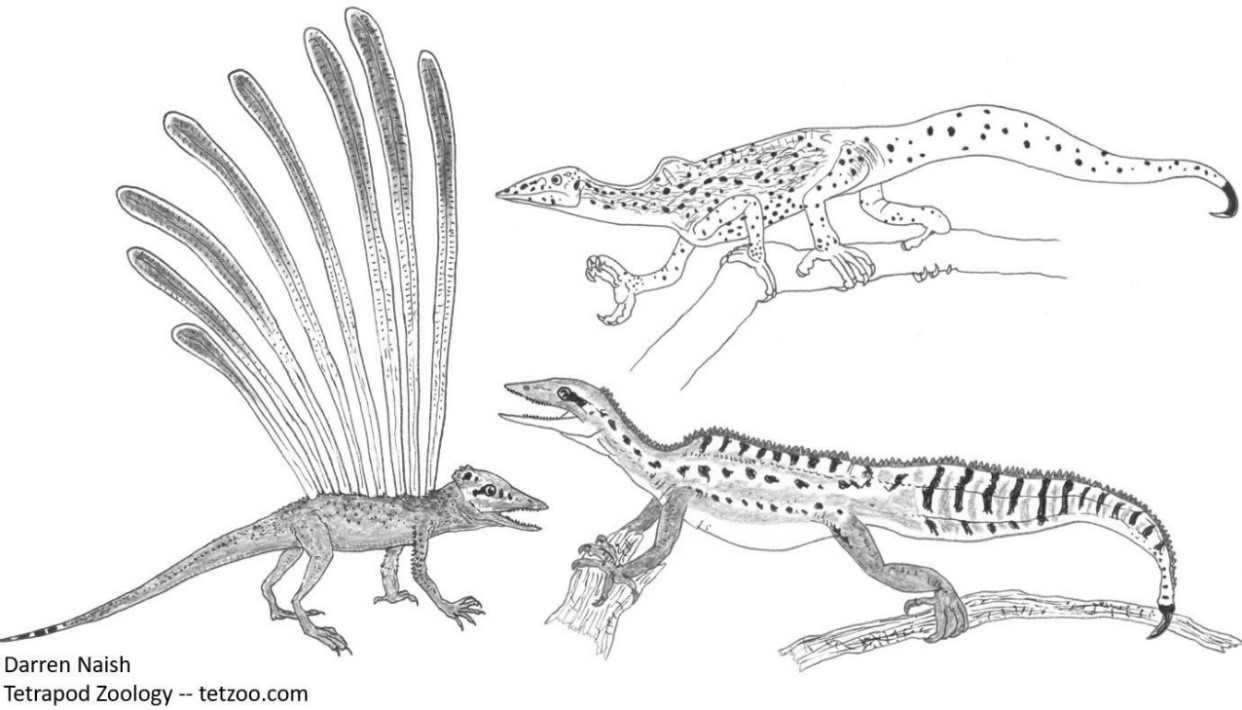
Longisquama (links) mit auffällig langen, blattähnlichen Körperanhängseln5) und die Drepanosauriden (rechts) sind kleine vierfüßige, kletternde, diapside Reptilien aus der Trias. Sie sind weder Archosaurier noch deren nahe Verwandte. Es gibt daher keinen Grund anzunehmen, dass sie für die Abstammung der Vögel relevant seien. FEDUCCIA und dessen Kollegen haben jedoch wiederholt auf die Möglichkeit verwiesen, dass diese Tiere näher mit den Vögeln verwandt seien als die (Nichtvogel-) Theropoden-Dinosaurier. Bild: Darren NAISH.
FEDUCCIA und die Debatte über die Halshaltung bei Sauropoden
Das wohl aufschlussreichste Kapitel des Buches ist das über die Halshaltung der Sauropoden, und mit "aufschlussreich" meine ich FEDUCCIAs Sicht auf die Wirbeltierpaläontologie, nicht die eigentliche Diskussion.
Die Fachwelt ist bezüglich der Halshaltung der Sauropoden in zwei Lager gespalten. Lager 1 argumentiert mit Hilfe digitaler Modellierung, dass Sauropoden auf eine halb-horizontale Halshaltung beschränkt waren und dass die Beweglichkeit des Halses aufgrund der überlappenden Facetten zwischen den Wirbeln relativ eingeschränkt war. Lager 2 argumentiert, Röntgendaten von lebenden Tieren würden darauf hindeuten, dass die Tiere eine eher vertikale Nackenhaltung einnahmen. Experimentelle Beweise würden zeigen, dass die überlappenden Facetten mehr Beweglichkeit zuließen, als von den fraglichen digitalen Modellen angenommen wird. Es wird darauf verwiesen, dass die digitale Modellierung eine Zuverlässigkeit der Rekonstruktion der Nackenanatomie vorspiegele, welche die (mehr oder weniger deformierten) Fundstücke nicht hergeben.
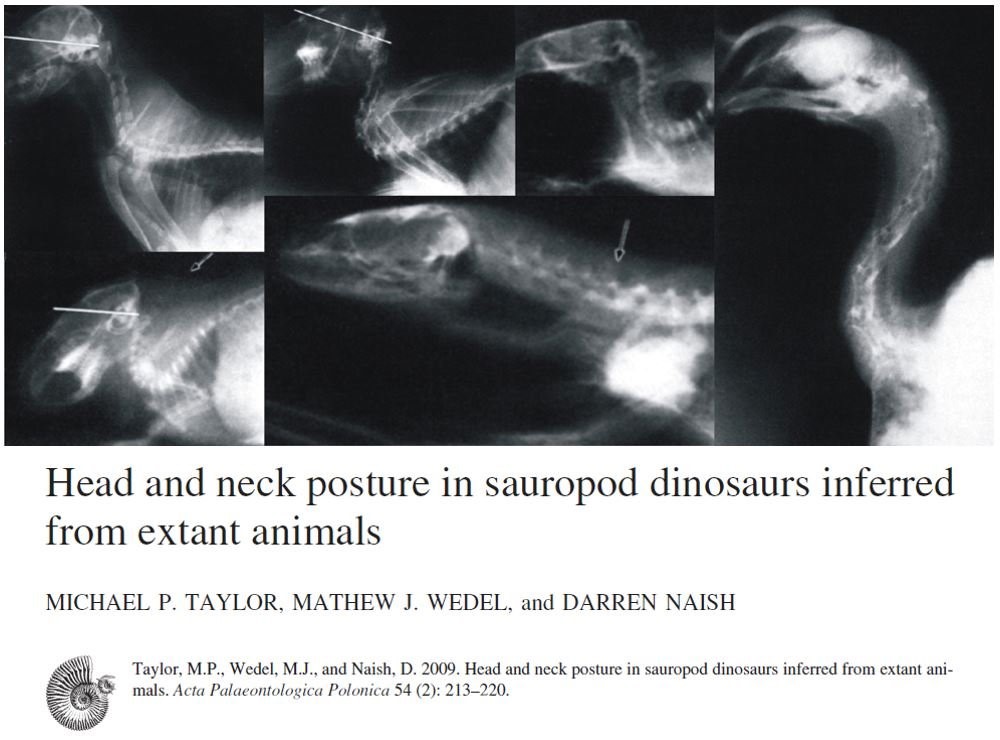
TAYLOR et al. (2009) argumentieren, die von lebenden Tetrapoden wie Amphibien, Säugetieren, Schuppenkriechtieren, Krokodilen und Vögeln gewonnen Daten würden nahelegen, dass eine eher vertikale Nackenhaltung für ausgestorbene Mitglieder dieser Gruppe angenommen werden müsse, zumal stichhaltige Belege für das Gegenteil fehlen.
Zur Klarstellung, ich reihe mich in das zweite Lager ein. FEDUCCIA zählt sich zum ersten Lager und bezeichnet die Angehörige von Lager 2 als "Antagonisten" (S. 37). Liegt das daran, dass er methodische Gründe kennt, warum die Argumente des zweiten Lagers abgelehnt werden sollten? Keineswegs. Er unterstellt, die Schlussfolgerungen des zweiten Lagers seien der ästhetischen Affinität gegenüber Abbildungen geschuldet, die Sauropoden gern mit Schwanenhälsen darstellen (S. 32-33).
Bei der Erörterung der Position von Lager 2 zitiert FEDUCCIA nur eine einzige relevante Arbeit (TAYLOR et al. 2009) und tut die darin enthaltenen Daten mit der Behauptung ab, sie würden nur von Haussäugetieren stammen, was unzutreffend ist. Er behauptet auch, dass Sauropoden in Bezug auf die Halsflexibilität nicht schwanenähnlich gewesen wären, aber das behauptet ohnehin niemand; es handelt sich dabei also um ein Strohmannargument.
Warum ist dieses Beispiel so aufschlussreich? FEDUCCIA scheint das Argument der halb-horizontalen Halshaltung als die traditionelle und biologisch sinnvollere Sichtweise zu betrachten. Doch Argumente wie die Nackenhaltung bei Sauropoden sind komplex; zahlreiche Autoren haben verschiedene Ansätze zu dem Thema gewählt. Zu unterstellen, dass eine bestimmte Sicht hierzu rückwärtsgewandt sei und rein aus ästhetischen Gründen bevorzugt werde, ist unredlich. Hier bedient sich FEDUCCIA erneut des Strohmannarguments und des subjektiven Unglaubens als Hauptwaffe.
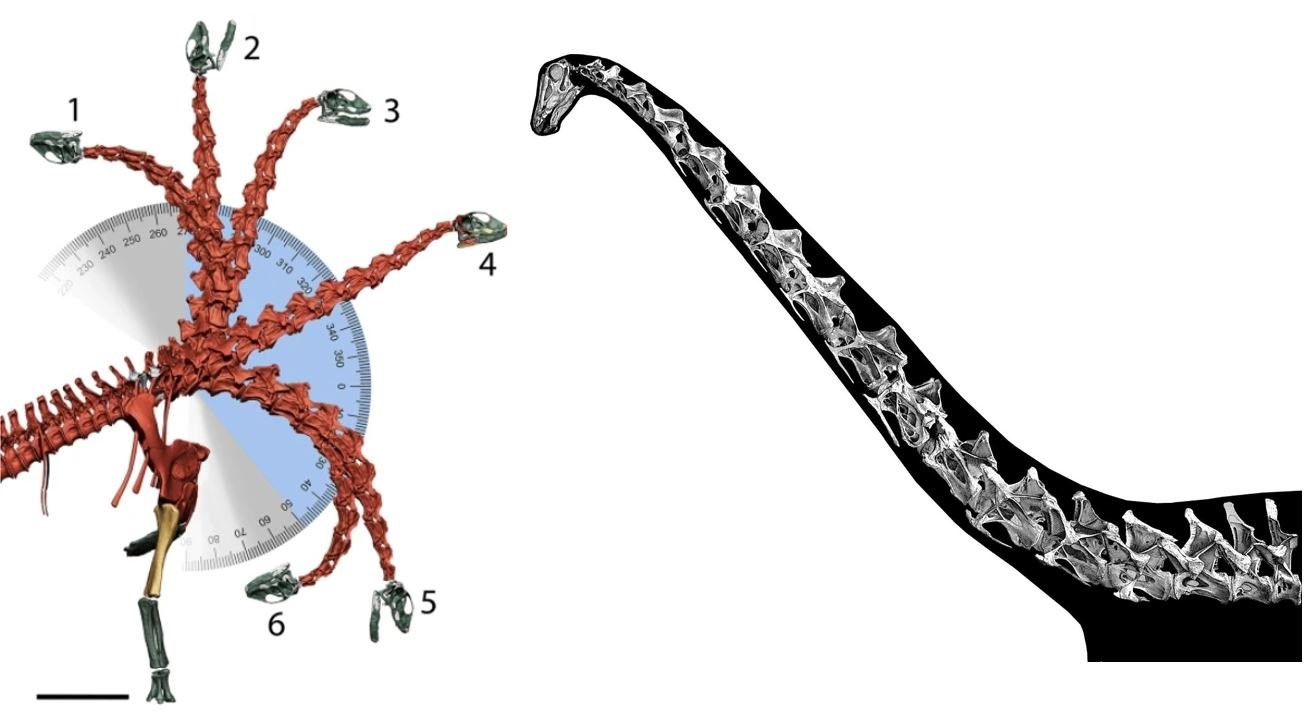
FEDUCCIAs Diskussion über Sauropoden soll den Leser glauben machen, dass die Hypothese des "aufrechten Halses" schlecht begründet sei und von Fundamentalisten vertreten werde, die eine konventionellere Sichtweise umstoßen wollen. Interessant ist dabei, dass FEDUCCIA die Arbeiten, die zu diesem Thema veröffentlicht wurden, vollkommen unzureichend bespricht. Das Bild links zeigt die postulierte Halsbeweglichkeit bei Spinophorosaurus, aus VIDAL et al. (2020a). Das Bild rechts zeigt Diplodocus in der gewohnten Nackenhaltung, die TAYLOR et al. (2009) aus den Daten lebender Tetrapoden ableiteten.
Interessanterweise hat sich viel getan, seit beide Lager ihre ersten Argumente austauschten. Inzwischen sind Studien zur biomechanischen Verteilung und Ableitung der Belastung (CHRISTIAN & DZEMSKI 2011), zur Ausrichtung der Wirbelsäule als Ganzes (VIDAL et al. 2020a) und vergleichende Studien zwischen Giraffen- und Sauropoden-Wirbelsäulenbereichen und -beweglichkeiten (VIDAL et al. 2020b) verfügbar. Sie kommen zu dem Schluss, dass Sauropoden eine eher vertikale Nackenhaltung einnahmen. FEDUCCIA zitiert oder erwähnt keine dieser Arbeiten, was seiner Gewohnheit entspricht Belege zu ignorieren, die den von ihm vertretenen Positionen widersprechen.
FEDUCCIA und die phylogenetische Systematik
Wenn FEDUCCIA die Phylogenie der Archosaurier erörtert, kommen mit schöner Regelmäßigkeit intellektuelle Unredlichkeit und naive Widerlegungsversuche ins Spiel. Sein Ziel ist es zu zeigen, dass das gesamte Feld chaotisch und unsicher sei und dass die taxonomischen Gruppen künstlich und schlecht definiert seien. So beginnt Kapitel 7 ("Dinosaurier: Was ist das?") mit einer Anekdote, in der FEDUCCIA einen namentlich nicht genannten (!) Kollegen bittet, den Begriff Theropode zu definieren. Besagter Kollege soll die tiefschürfende Antwort gegeben haben: "Ein Theropode ist, was immer diese Leute über diesen sagen!" (S. 93).
[MN & AB: FEDUCCIA scheint nicht zu interessieren, dass die Theropoden eine seit 140 Jahren anerkannte Untergruppe der Dinosaurier sind, die unzählige Male anhand apomorpher Merkmale definiert worden ist, in neuerer Zeit beispielsweise von GAUTHIER (1986), SERENO (1999), RAUHUT (2003), NESBITT et al. (2009), CAU (2018) und vielen anderen.]
FEDUCCIA fährt fort, dass Dinosaurier durch "kaum andere morphologische Merkmale als die Größe und den Gang" (S. 94) definiert worden seien und dass solche Merkmale, die zur Definition von Theropoden verwendet werden, die Folge einer konvergenten Evolution der Zweibeinigkeit sein könnten. Schließlich wisse man, dass bestimmte Nicht-Dinosaurier-Archosaurier der Trias oberflächlich gesehen Theropoden ähnelten. Nebenbei bemerkt er, dass bestimmte Theropoden-Merkmale nicht bei allen Theropoden vorhanden seien, was ein Problem darstelle. Das ist entweder bemerkenswert naiv oder besonders trickreich: Die Evolution bedingt doch gerade, dass nicht alle Mitglieder einer Abstammungsgruppe Merkmale zeigen, die bei den frühesten Vertretern dieser Gruppe noch vorhanden waren.
Ein philosophisches Problem, das bei FEDUCCIA immer wieder auftaucht, ist sein Beharren darauf, dass die Gruppenzugehörigkeit in der Phylogenetik anhand einiger weniger, offensichtlicher, anatomischer Merkmale nachgewiesen werden sollte. Organismen sind jedoch komplexe Entitäten - und wenn die Phylogenetik uns eines lehrt, dann, dass eine große Menge an Daten analysiert werden muss, wenn wir eine Phylogenie ermitteln wollen, die den wahren Verwandtschaftsverhältnissen möglichst nahekommt.
Die Identifikation von zwei oder drei "Schlüsselmerkmalen" ist vollkommen unzureichend. Es ist erschreckend naiv anzunehmen, es sei gerechtfertigt, ein oberflächlich dinosaurierähnliches Reptil aufgrund des Besitzes einer langen Schnauze und spitzer Zähne als Dinosaurier zu charakterisieren, angesichts dutzender anatomischer Details, die ihm einen ganz anderen Platz im Kladogramm zuweisen.
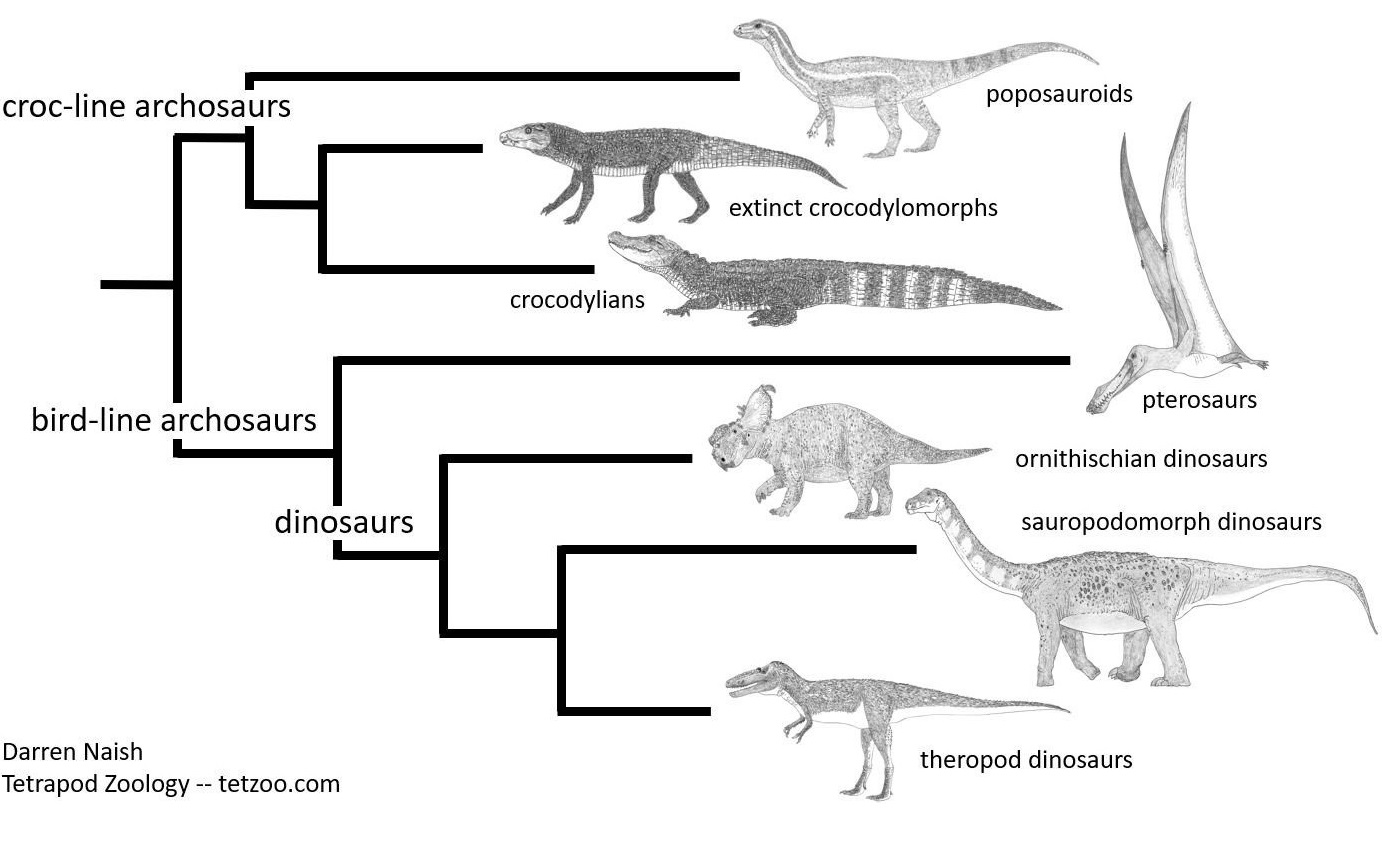
Grob vereinfachte Darstellung der Archosaurier-Evolution, wie sie derzeit diskutiert wird. Eine Reihe von Arten auf der Stammlinie der Krokodile, wie etwa der Poposauroide Effigia ganz oben im Diagramm, ähneln oberflächlich bestimmten Dinosauriern. Eine Fülle anatomischer Details belegt aber, dass sie nicht zur Stammlinie der Vögel gehören und mit Dinosauriern nichts zu tun haben. Das hier dargestellte Kladogramm stützt sich auf eine Vielzahl von Studien, die eine enorme Anzahl anatomischer Beobachtungen und Entdeckungen beinhalten. Bild: Darren NAISH.
FEDUCCIA und die neuzeitlichen Vögel (Neornithes)
Kommen wir zu FEDUCCIAs Ausführungen über die Evolution der Neornithes, einem Feld, das FEDUCCIAs Kritiker in der Regel meiden. FEDUCCIA versucht mit diesem Thema zu punkten; er vertritt die in seinen Augen bahnbrechende Vorstellung, dass nach der Kreidezeit eine Art explosive Radiation oder "Urknall" der Vogelevolution stattgefunden habe. Er behauptet, moderne Erkenntnisse würden seine Idee stützen. Das kann ich nicht nachvollziehen. FEDUCCIA hat zwar Recht damit, dass die Radiation ihren Höhepunkt nicht schon früh in der Kreidezeit erreichte. Ja, die Hauptradiation der Vögel ereignete sich in der Tat erst nach der Kreidezeit. Funde wie der kürzlich beschriebene Asteriornis, ein enger Verwandter der heutigen Hühner und Gänse, belegen aber, dass sie schon lange vor dem Ende der Kreidezeit begann. Die neuesten Arbeiten zeigen, dass sich die wichtigsten Gruppen (Urkiefervögel, Hühner- und Gänsevögel sowie Neoaves) bereits vor 100 bis 75 Mio. Jahren auseinander entwickelten (z.B. FIELD et al. 2020).
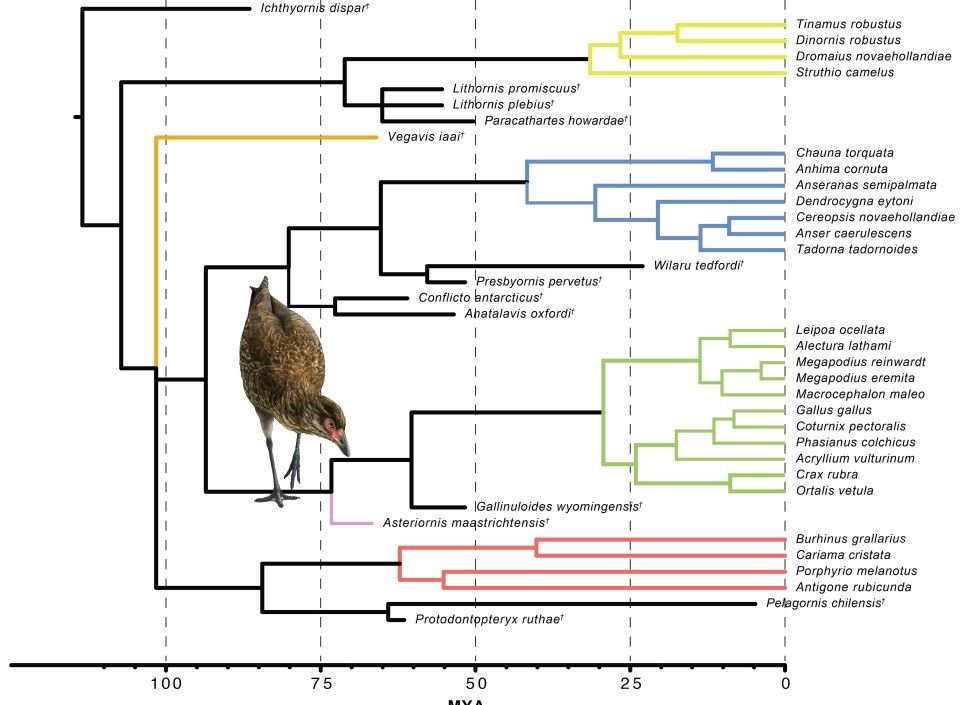
Heute unterstützen sowohl der Fossilienbefund als auch die zeitlich kalibrierten molekularen Stammbäume das gleiche Szenario: Die neuzeitlichen Vögel entstanden während der späten Kreidezeit, allerdings nicht erst an deren Ende. Dieses Kladogramm zeigt, dass sich die Hauptkladen der neuzeitlichen Vögel bereits vor rund 100 Millionen Jahren begannen auseinander zu entwickeln. Am Ende der Kreidezeit hatten sich schon diverse Abstammungslinien etabliert. Unbestritten ist freilich, dass der größte Teil erst nach der Kreidezeit entstand. Aus FIELD et al. (2020).
Während FEDUCCIAs eigene Diagramme zeigen, dass die Neornithes aus der späten Kreide stammen (S. 297), bestreitet er in seinem Text die Existenz neuzeitlicher Vögel in der späten Kreidezeit. Er zeichnet ein verwirrendes Bild, und ich finde es unredlich, wenn er behauptet, damit etwas Wertvolles zur Diskussion beigesteuert zu haben.
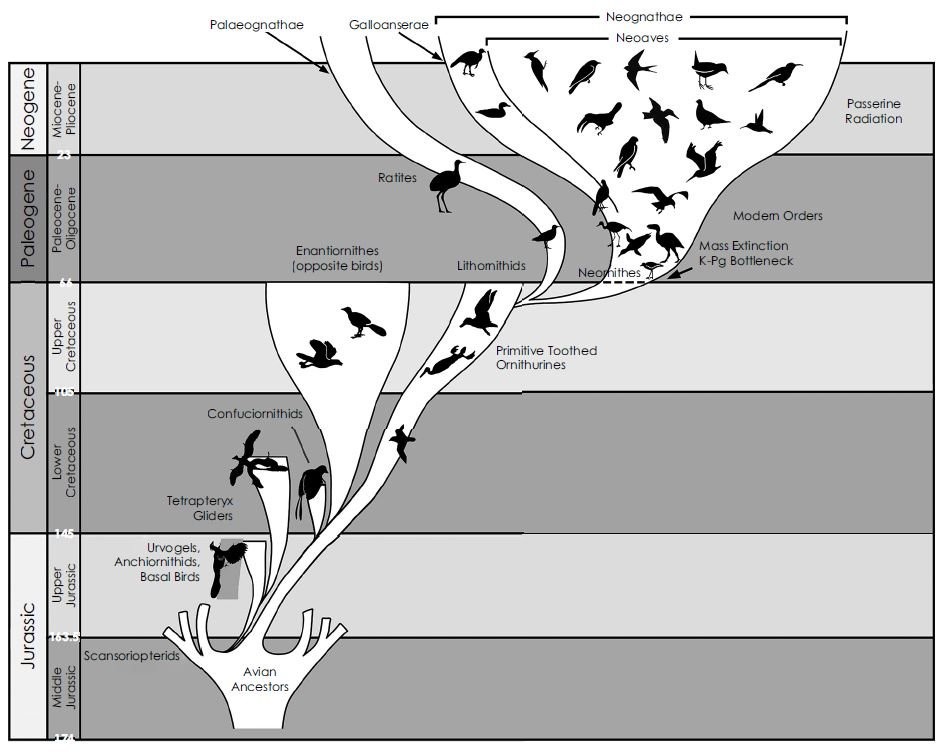
Die in RTBAD dargestellte Version vom "Urknall" der neuzeitlichen Vögel. Sie unterscheidet sich von den Versionen, die FEDUCCIA früher veröffentlichte, insoweit, als sie den Ursprung der Neornithes-Radiation in die späte Kreidezeit zurückverlegt. Bild: FEDUCCIA (2020).
Ein weiteres Beispiel für Strohmänner: FEDUCCIA greift die phylogenetische Systematik aufgrund der Tatsache an, dass sich die Zuordnung einiger Taxa im Kladogramm immer wieder ändert. Seiner Meinung nach beweise die Korrektur einer Klade bestehend aus Haubentauchern und Seetauchern, dass die phylogenetische Systematik nicht funktioniere.
Nochmals: Ja, in der Wissenschaft werden Fehler gemacht, und die Begründung einer Haubentaucher-Seetaucher-Klade war ein solcher. Der Grund war die Verwendung unzureichenden Datenmaterials. Es ist unfair, dies als eine Art Rechtfertigung dafür zu benutzen, die Hände in den Schoß zu legen und mit der Wissenschaft aufzuhören. Es ist ja nicht so, dass FEDUCCIAs Bemühungen zur Rekonstruktion verwandtschaftlicher Beziehungen zwischen Organismen qualitativ bessere Resultate erbrächten. Falsche phylogenetische Hypothesen erstellte FEDUCCIA in seiner eigenen Forschung zuhauf (FEDUCCIA 1977a, 1977b, 1978, OLSON & FEDUCCIA 1980a, 1980b).

Es stimmt, dass Joel CRACRAFT einst Seetaucher (oben links), Lappentaucher wie den Haubentaucher (oben rechts) und die Hesperornithinen (unten) in der Klade der "Gaviomorphae" zusammenfasste. Andere Autoren bildeten in jüngerer Zeit eine Klade aus Seetauchern und Haubentauchern. Beweisen solche Irrtümer, dass Versuche, die Stammesgeschichte der Vögel mit Hilfe der Kladistik zu rekonstruieren, zum Scheitern verurteilt und daher Zeitverschwendung sind? Nein, sie beweisen, dass die Wissenschaft fehleranfällig ist, aus Fehlern lernt und sich so allmählich "empor irrt". Bilder: Seetaucher von John Picken from Chicago, USA, Gavia immer -Minocqua, Wisconsin, USA -swimming-8, Bild leicht beschnitten, CC BY 2.0. Die übrigen Bilder stammen von Darren NAISH.
{kind=link}
In ähnlicher Weise versucht FEDUCCIA die Leser in die Irre zu führen, indem er eine Litanei an Beispielen phylogenetischer Hypothesen herbetet, die heute als widerlegt gelten. Zu den Beispielen, die er anführt, gehören die einst beliebte Verknüpfung von Walen mit Mesonychiern, die Einordnung der Dromornithiden als Laufvögel und die vorgeschlagene Verwandtschaft zwischen Ruder- und Lappenenten. FEDUCCIA will damit demonstrieren, dass die phylogenetische Standard-Systematik drastische Mängel aufweise. Aber noch einmal: Das ist nicht das, was diese Beispiele aufzeigen. Sie zeigen, dass Phylogenetiker Fehler machen, wenn sie es versäumen, relevante Daten zu berücksichtigen (oder, noch besser, Daten, die noch gar nicht bekannt waren, als die betreffende Hypothese aufgestellt wurde). Dies ist kein fataler Fehler, sondern die unvermeidliche Folge wissenschaftlichen Fortschritts.
FEDUCCIA stellt ferner die Behauptung auf, dass die Flugsaurier einst als Schwestergruppe der Theropoden betrachtet wurden. In Wahrheit wurde lediglich behauptet, dass Flugsaurier eng mit den Dinosauriern verwandt seien, was noch immer zutrifft. FEDUCCIA verwechselt dies mit der Hypothese, dass Flugsaurier früher als "obligat zweibeinig" galten und vom Boden aus starteten. Zwar hängt diese Hypothese mit dem Argument zusammen, dass Flugsaurier eng mit Dinosauriern verwandt sind; sie wurde aber als Argument der funktionellen Anatomie vorgebracht und ist daher in FEDUCCIAs Liste umstrittener phylogenetischer Hypothesen fehl am Platz.
Einen weiteren Strohmann präsentiert FEDUCCIA mit der Behauptung, dass Ornithologen weithin und unkritisch die altmodische Ansicht vertraten, dass Laufvögel ihre Verbreitung der Vikarianz verdankten.6) Diese Ansicht wurde in den 1980er Jahren von Joel CRACRAFT und von einigen Wissenschaftsautoren vertreten (FEDUCCIA zitiert hier Richard DAWKINS). Aber wird diese Hypothese heute noch akzeptiert? Keineswegs. Es ist irreführend zu behaupten, dass sie heute noch gälte.
Bei den Laufvögeln und ihren Verwandten vertritt FEDUCCIA die Ansicht, dass diese Vögel ihre Anatomie der Pädomorphose verdanken. [MN & AB: Das bedeutet, Jugendstadien eines Vorfahren bleiben im Erwachsenenstadium erhalten]. Das war zwar früher eine populäre Hypothese, aber es spricht heute viel dagegen; siehe den Artikel Controversies in Ratite and Tinamou Evolution (Part I) in Tetrapod Zoology. Aus FEDUCCIAs Perspektive würde man das nicht erkennen, denn er vertritt voller Überzeugung den Standpunkt, dass eine pädomorphe Erklärung für die Morphologie der Laufvögel so selbstverständlich sei, dass sie nicht in Frage gestellt werden sollte. Immer wieder wirkt die Art, wie FEDUCCIA ein Thema behandelt, irreführend; er gibt die von den Fachleuten vertretene Position häufig falsch wieder.
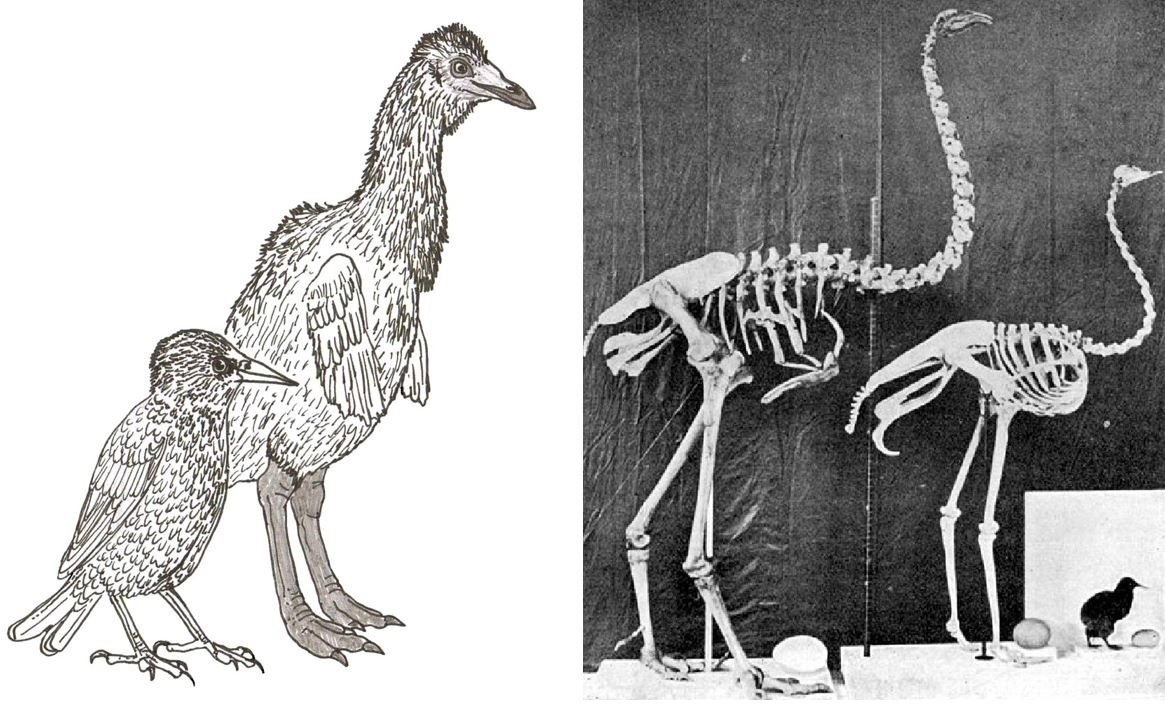
Die Hypothese, Laufvögel seien pädomorph, ist weit verbreitet. Ob das stimmt, wissen wir nicht. Die Hypothese scheint nicht so gut zu den Daten zu passen. Bilder: Darren NAISH.
Die endlose Debatte über die Homologie der Vogelfinger
In den letzten Jahren beschäftigte sich FEDUCCIA häufig mit der Homologie der Vogelhand. Er argumentiert, dass die Fingerzählweise bei Vögeln und Theropoden unterschiedlich sei und dass die Vögel daher nicht den Theropoden zugerechnet werden könnten. Darüber hinaus behauptet er, seine Arbeiten dienten dazu, "Licht in diese Angelegenheit zu bringen" (S. 138). Ich argumentiere mal zynisch; ich denke, dass diese Arbeiten speziell in der Absicht erstellt wurden, die phylogenetische Verbindung zwischen Vögeln und Dinosauriern zu kappen (siehe dazu meine obenstehenden Ausführungen zur Bestätigungsverzerrung).
FEDUCCIA unterstellt, die Fachwelt würde die Finger der Vögel und die ihrer mutmaßlichen Dinosauriervorfahren nachträglich für homolog, das heißt die scheinbar widersprüchlichen Fingerformeln7) für identisch erklären, nur um die Daten willkürlich an ihr Kladogramm anzupassen (S. 142). Fair ist das nicht, denn die Homologie der Finger lässt sich aus der gesamten Phylogenie ableiten und ist unabhängig davon, ob man Kladogramme verwendet oder nicht.
[MN & AB: Zum Beispiel belegen Fossilienreihen die Reduktion von zwei Fingern an der Handaußenseite unter den Vorfahren von Deinonychus. Die übrig gebliebenen Finger ähneln den drei Fingern von Archaeopteryx.]
Wer den Konsens der Fachwelt zur Entwicklung der Vogelhand nachzuvollziehen versucht, wird feststellen, dass mehrere Modelle diskutiert werden. Dabei ist wichtig zu betonen, dass selbst Wissenschaftler, die wie FEDUCCIA die Indexierung der Vogelfinger als Zeige-, Mittel- und Ringfinger (II-III-IV) favorisieren (ČAPEK et al. 2013, DE BAKKER et al. 2021), keine Unvereinbarkeit mit der Theropoden-Abstammung feststellen.
Zudem ist die Datenlage nicht eindeutig: Einige Studien zur Expression der Hox-Gene stützen die I-II-III-Konfiguration (VARGAS & FALLON 2005, SALINAS-SAAVEDRA et al. 2014), aber nicht alle (STEWART et al. 2019, DE BAKKER et al. 2021). Angesichts der widersprüchlichen Befunde ist es amüsant zu sehen, wie sich FEDUCCIA verbiegt, um unliebsame Studien weg zu erklären: Nur, weil etwas experimentell gezeigt werden kann, bedeute das nicht, dass es in der Natur auch so passiert sei (S. 144).
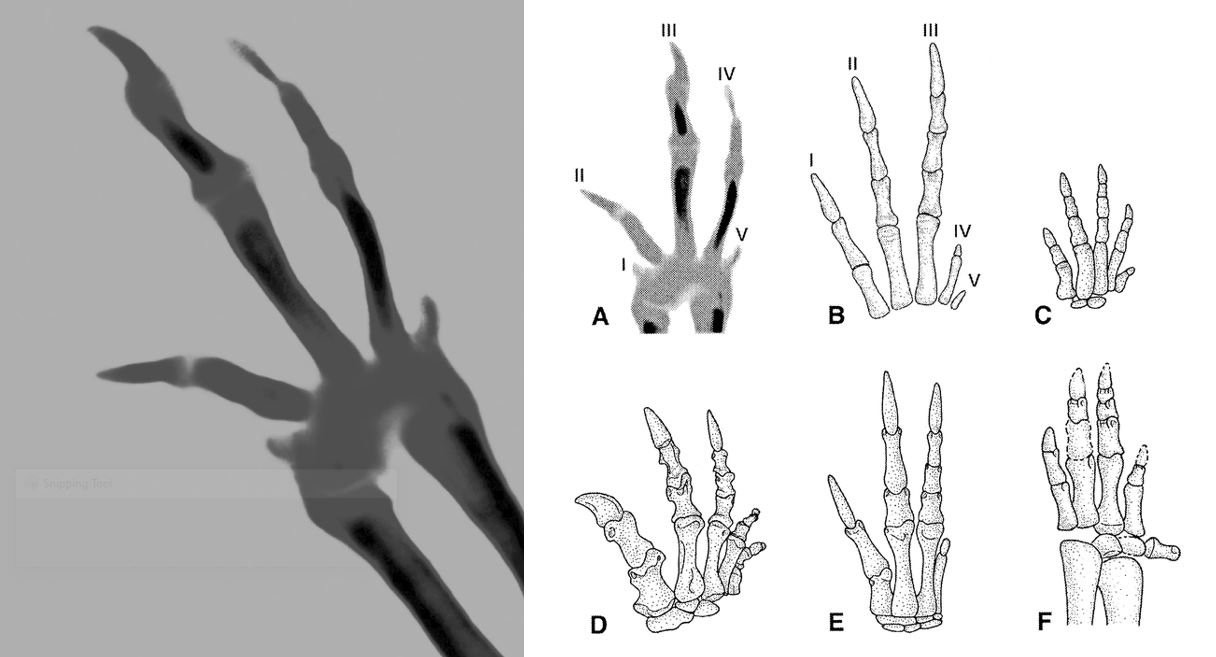
Links: die rechte Hand eines Straußenembryos in Dorsalansicht. Rechts: die gleiche Hand (A) im Vergleich zu der von Nichtvogel-Dinosauriern (B bis F). Ein wesentlicher Bestandteil von FEDUCCIAs Identifizierung der Fingerzählweise in der Vogelhand stützt sich auf das Argument, dass Straußenembryonen fünf Finger besäßen. Die kleinen "Knospen" an der Mittelhand würden die Finger Nummer I und V repräsentieren. VARGAS & FALLON (2005, S. 214-215) betonen jedoch, dass diese "Verdichtungen am Flügel in einem viel späteren Stadium auftreten als die Fingerstrahlen, darüber hinaus sehr klein und nur vorübergehend nachweisbar sind, sodass diese mesenchymalen Verdichtungen nicht mit Sicherheit mit bestimmten Fingern identifiziert werden können, die im Erwachsenenalter verloren gehen". FEDUCCIA (2020, S. 151) zitiert Henry GEE mit den Worten: "Ich kann mit diesem Bild nicht viel anfangen; es erinnert mich an einen Rorschach-Test". Abbildungen: Links: FEDUCCIA (2020). Rechts: FEDUCCIA (2002).
Um die Fingerformel (II-III-IV) bei den Vögeln mit der (I-II-III)-Indexierung der Finger bei ihren tridaktylen Vorfahren in Einklang zu bringen, haben einige Autoren eine Frameshift [MN & AB: eine Verschiebung des genetischen Bezugssystems] vorgeschlagen. Das bedeutet, dass Zeige-, Mittel- und Ringfinger der Vögel in der Embryonalentwicklung den zur Verfügung stehenden Platz der reduzierten übrigen Finger ausnutzen und weiter nach innen wachsen. Dies führt dazu, dass die biomechanisch verlagerten Finger (II-III-IV) unter den Einfluss veränderter Signalfaktor-Konzentrationen geraten und das Genexpressionsmuster der inneren Finger (I-II-III) annehmen.
Diese als Frameshift bezeichnete Hypothese ist entwicklungsbiologisch sehr interessant. FEDUCCIA weist das von der Hand und fragt nach Sinn und Grund dafür (S. 144). Später argumentiert er, die Frameshift sei unsinnig, da sie keine Funktion erfülle.8) Es sollte aber klar sein, dass diese Hypothese eine Reihe überraschender Befunde erklären kann. FEDUCCIA mag zwar mehr an Vögeln als an anderen Theropoden interessiert sein. Tatsache ist aber, dass auch Nichtvogel-Theropoden bemerkenswerte Hände haben; das Vorhandensein von Didaktyl- und sogar Monodaktyl-Linien bedeutet, dass wir bezüglich der Handevolution der Theropoden auch dann von einer Frameshift sprechen könnten, wenn es keine Vögel gäbe.
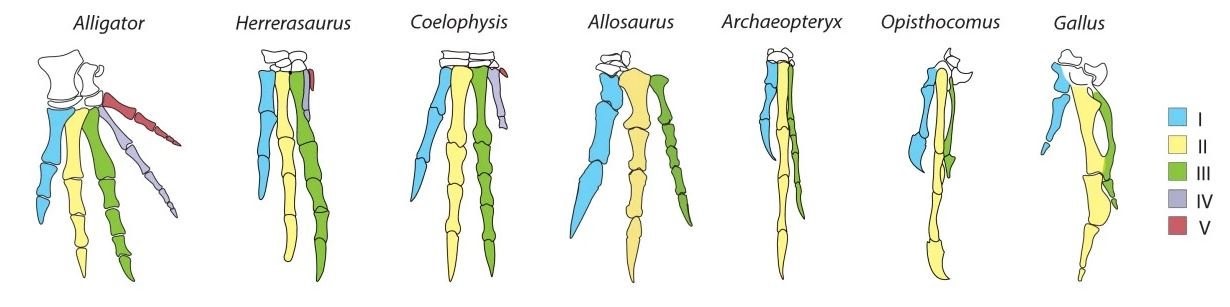
Ausgehend vom Muster der Handanatomie in der Evolutionsgeschichte scheint es offensichtlich, dass die Vogelhand aus den Fingern I, II und III besteht. Doch der Schein kann trügen. Was sagen die molekularen Daten? Durch die Untersuchung der Expression von HoxD-Genen in der sich entwickelnden Hand stützen einige Studien die I-II-III-Konfiguration. Bild: SALINAS-SAAVEDRA et al. (2014).
Was die "Funktion" betrifft, so wissen wir nicht, warum diese Ereignisse eintraten. Ein vernünftiges Gegenargument ist das aber nicht. Die Tatsache, dass auch bei Salamandern und Glattechsen über Frameshift berichtet wurde (wogegen FEDUCCIA andeutet, das Phänomen der Frameshift sei außerhalb des vorgeschlagenen Theropoden-bzw. Vogel-Beispiels gar nicht bekannt), ist für die Funktion und Struktur dieser Tiere nicht bedeutsamer als theoretisch für Vögel und andere Theropoden.
Das "Kollagenmodell" für Dino-Flaum ist schlechte Wissenschaft
Eine weitere Debatte, in der FEDUCCIA offenbar einen unverrückbaren, zur Meinung der Fachwelt konträren Standpunkt vertritt, betrifft die aus verschiedenen Dinosaurier- und Flugsaurier-Fossilien bekannten Integument-Fasern bzw. Filamente.
[MN & AB: Dabei handelt es sich um haarartige Keratin-Fasern, die morphologisch und mikrostrukturell deutliche Ähnlichkeiten mit Federn aufweisen.]
FEDUCCIA ist bemüht, seine Leser glauben zu machen, dass es sich bei der flaumartigen Körperbedeckung in Wahrheit um Kollagenfasern der Haut handele. Diese Hypothese wurde schon früh vorgeschlagen, nämlich als eine Welle von Entdeckungen gefiederter Dinosaurier und Flugsaurier vorrangig aus China begann. Man kann sich daher nur schwer des Eindrucks erwehren, dass FEDUCCIA die Hautanhängsel als Kollagenfasern interpretiert sehen möchte, nur um dem Eindruck entgegen zu wirken, Dinosaurier und Flugsaurier seien irgendwie vogelähnlich gewesen.
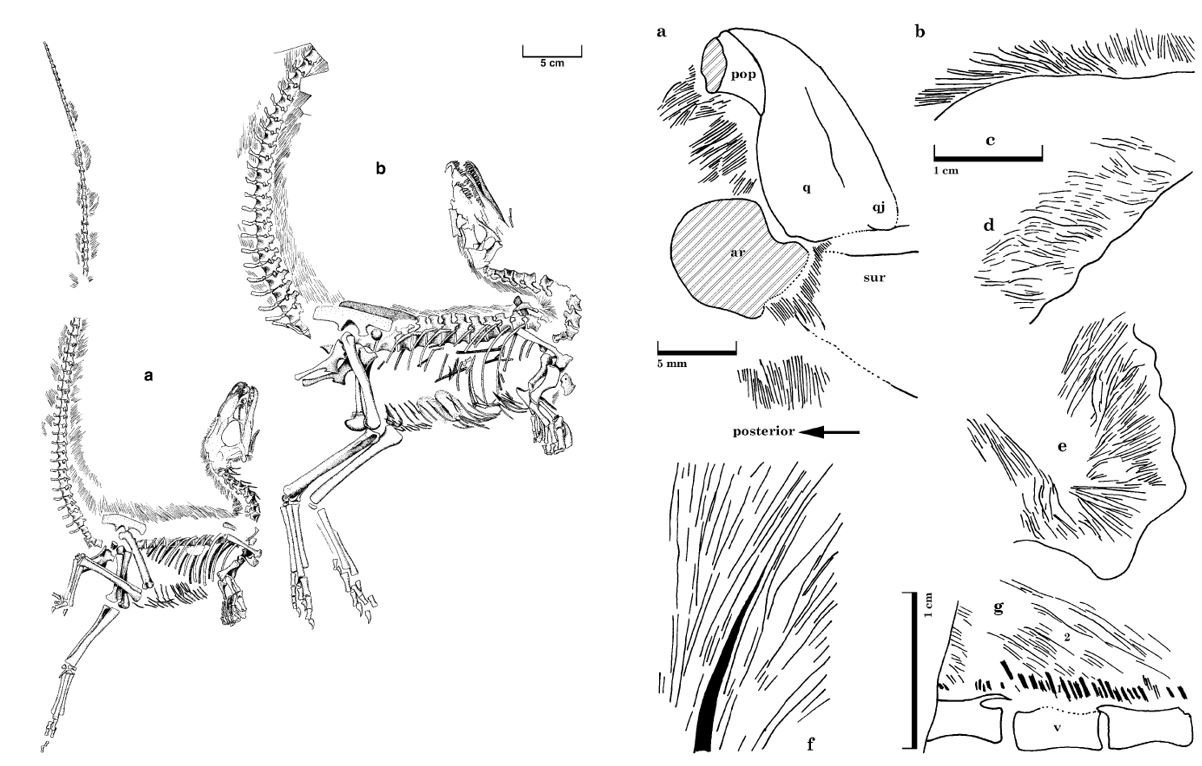
In der Diskussion über den Ursprung der Vögel wird von einigen die Meinung vertreten, dass man dem Dinosaurier-"Flaum" gegenüber sehr skeptisch sein sollte. Skepsis ist in dieser Frage tatsächlich angebracht. Aber regelrechte Verleumdungen auszusprechen, wie es FEDUCCIA tat (MORELL 1997, S. 38), war und ist ein Zeichen massiver Voreingenommenheit. Die Bilder zeigen zwei Sinosauropteryx-Exemplare mit fadenförmigen Integumentstrukturen [(a) NIGP 127586 und (b) NIGP 127587] sowie Nahaufnahmen dieser Filamente, die am gesamten Körper erhalten sind (und nicht nur an der dorsalen Mittellinie, wie FEDUCCIA und Kollegen behaupten). Bilder: CURRIE & CHEN (2001).
In RTBAD behauptet FEDUCCIA, solide Wissenschaft habe die Hautfilamente als Kollagenfasern identifiziert. Aber selbst die großzügigste Auslegung der Daten zeigt, dass dies nicht zutrifft. Es handelt sich bestenfalls um eine spekulative Hypothese. "Weshalb weigern sich die Paläontologen, die Möglichkeit in Betracht zu ziehen, dass diese Filamente Kollagenfasern darstellen?" (S. 184), fragt FEDUCCIA. Erstens, weil beiderlei Strukturen verschieden aussehen. Zweitens, weil der Dino-Flaum Merkmale zeigt, die darauf hindeuten, dass er aus Keratin und nicht aus Kollagen besteht. Und drittens, weil diejenigen, die andere davon überzeugen wollten, dass die Hautanhängsel aus Kollagen bestehen, sich sehr angestrengt und es vermasselt haben (man könnte auch sagen: geschummelt), um Kollagenfasern wie Dinosaurierflaum aussehen zu lassen.
Ich sage "geschummelt", weil FEDUCCIA anatomische Präparate abbildet, bei denen Kollagenfasern aus dem Knochen herausgelöst wurden, um Objekte zu erzeugen, die den Integument-Filamenten ähneln. Man darf zurecht fragen, welche Bedeutung solche Laborpräparate für fossile Dinosaurier und Flugsaurier mit wuscheligem Fell haben sollen. Um es mit FEDUCCIA zu formulieren: Nur "weil etwas experimentell gezeigt werden kann, heißt das nicht, dass es in der Natur vorkam" (S. 144).

Wesentlich für FEDUCCIAs Interpretation, dass es sich bei den Filamenten des Sinosauropteryx um Kollagenfasern handele, ist die Deutung, dass sich die Filamente "unterhalb der Haut" (S. 185) bzw. innerhalb der "klar abgegrenzten Körperkonturen" (S. 165) befänden. Bei dem hier gezeigten Exemplar eines Sinosauropteryx (IVPP V12415) (das Bild zeigt einen Teil des Schwanzes) wurde die vermeintliche Haut- bzw. Körperkontur mit roten Pfeilen markiert. Tatsächlich aber handelt sich hier mitnichten um den Körperumriss, sondern um ein Präparations-Artefakt: Die Pfeile grenzen die Zone, in der die Gesteinsmatrix durch den Einsatz der Präparationswerkzeuge abblätterte, von dem Bereich ab, in dem die Matrix erhalten blieb. Dies ist bei all den betreffenden Exemplaren der Fall. Die meisten Beobachtungen, die zur Unterstützung der Kollagenfaser-Hypothese herangezogen werden, sind von diesem Kaliber. Bild: Darren NAISH.
Weiterhin bildet FEDUCCIA dünne, mehrfach verzweigte Bänder aus kollagenem Bindegewebe (sog. Ligamente) aus dem Bewegungsapparat der Vögel ab, um zu demonstrieren, dass sie den Hautanhängseln von Dinosauriern und Flugsauriern ähneln. Oberflächlich betrachtet mag das zutreffen. Aber woher stammen die Ligamente? Ragen sie wie der haarähnliche Flaum der Dinosaurier von der Außenseite des Körpers ab? Nein, es handelt sich um anatomische Präparate, die nur in bestimmten Körperregionen vorkommen, wie etwa zwischen den Dornfortsätzen der Halswirbel.
Der einzige Fachmann, der FEDUCCIAs Deutung der Hautanhängsel teilte, war der inzwischen verstorbene Paläontologe Theagarten LINGHAM-SOLIAR. LINGHAM-SOLIAR wird mehrfach in RTBAD erwähnt. Ein kurzer Exkurs über LINGHAM-SOLIAR ("Solly" für seine Freunde und Diskussionspartner, zu denen ich auch gehörte) lohnt sich: In den späten 1990er und frühen 2000er Jahren veröffentlichte LINGHAM-SOLIAR Arbeiten zur Beschaffenheit der Haut von Ichthyosauriern, wobei sein Hauptinteresse den komplexen, mehrschichtigen Kollagennetzen galt. Diese sind in die Haut eingebettet und bestehen aus sich überlappenden, gitterartigen Faserbündeln.
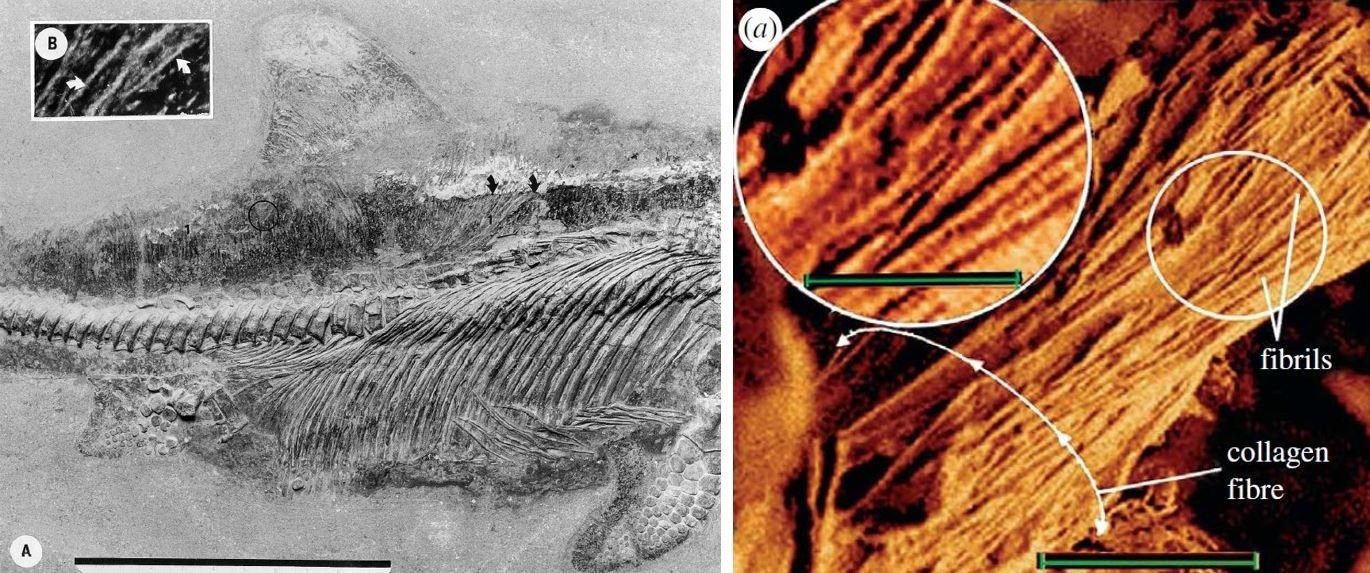
Links: Ichthyosaurier-Hautfasern. Das Foto zeigt, dass die Fasern weit innerhalb des äußeren Körperumrisses liegen, das heißt tief in der Haut eingebettet sind. Rechts eine Nahaufnahme von Ichthyosaurier-Hautfasern, wie sie von LINGHAM-SOLIAR & WESLEY-SMITH (2008) zur Verfügung gestellt wurde. Man beachte, dass sich die zahlreichen winzigen Fibrillen nicht zu ihren Enden hin verjüngen, sondern zu dicken, seilartigen Strukturen vereinigen. Strukturell haben sie somit keine Ähnlichkeit mit dem Dinosaurier-Flaum. Bilder: Links: aus LINGHAM-SOLIAR (2001), Maßstab: 10 cm. Rechts: LINGHAM-SOLIAR & WESLEY-SMITH (2008), Maßstab: 2 Mikrometer; hineingezoomter Bereich: 1 Mikrometer.
Die Struktur dieser Faserbündel sieht nicht aus wie die Hautanhängsel der Dinos und Flugsaurier. Leider war LINGHAM-SOLIAR anderer Meinung; er veröffentlichte zwischen 2007 und 2015 Arbeiten und Bücher, in denen er wie FEDUCCIA zu zeigen versuchte, dass es sich bei den Filamenten der Dinosaurier um verkannte Kollagenfasern handelt. Ich habe mich oft gefragt, ob LINGHAM-SOLIAR die Kollagenfasern der Ichthyosaurier erst als relevant für dieses Thema betrachtete, nachdem ihn FEDUCCIA mit der Nase darauf stieß. Wie also wurden LINGHAM-SOLIARs Argumente, abgesehen von FEDUCCIA, rezipiert? Die primäre Reaktion war Ablehnung, denn gründlichere Untersuchungen ergaben, dass es sich um nicht-kollagene Fasern handelt, sondern um solche mit einzigartiger, für Haare und Federn typischer mikroskopischer Anatomie (MAYR 2010, ZHANG et al. 2010, GODEFROIT et al. 2014, 2020, MAYR et al. 2016, SMITHWICK et al. 2017).
[MN & AB: ZHANG et al. (2010) wiesen sogar fossile Melanosomen (Organellen von Pigmentzellen) in den Filamenten von Sinosauropteryx nach, die Federn und Haaren Farbe verleihen. Die Partikel entsprachen in Größe, Anordnung und Verteilung den Ultrastrukturen fossiler Melanosomen, die in echten Vogelfedern identifiziert wurden. Kollagenfasern enthalten dagegen keine Melanosomen. Und würden sie aus der Haut stammen, würde man eine homogene Verteilung erwarten, was nicht der Fall ist. Im Gegenteil, anhand der Verteilung ließ sich nachweisen, dass der Schwanz von Sinosauropteryx eine hell-dunkel-gestreifte Bänderung aufwies. Die Deutung als dermale Kollagenfasern ist daher abwegig.]

Bilder oben: Bereiche mit außergewöhnlich gut erhaltenen haarartigen Hautanhängseln am Schwanz von Sinosauropteryx. Sie zeigen glatte, haarartige Filamente, die sich zu den Spitzen hin verjüngen, so wie dies für Keratin-Fasern (Haare, Federn) typisch ist, ohne Anzeichen von Wülsten, wie sie bei abgebautem Kollagen vorkämen. Bild unten links: Elektronenmikroskopische Aufnahme von Melanosomen, wie man sie in echten Federn des Vogels Confuciusornis fand. Unten rechts zum Vergleich Melanosomen, die in den Integument-Fasern von Sinosauropteryx gefunden wurden; die Ultrastrukturen sind identisch. Maßstab der elektronenoptischen Bilder: 2 Mikrometer. Bilder oben: aus SMITHWICK et al. (2017). Bilder unten: aus ZHANG et al. (2010). [MN & AB]
LINGHAM-SOLIARs Eindruck, dass der Dino-Flaum von Sinosauropteryx aus Kollagenfasern bestehe, ist teilweise der geringen Vergrößerung seiner Aufnahmen geschuldet, teils handelt es sich um falsch identifizierte Präparationsmarkierungen und Löcher in der Matrix. FEDUCCIA erwähnt diese Sachverhalte nicht. Hinsichtlich der Kritik von SMITHWICK et al. (2017) an LINGHAM-SOLIAR schreibt er, ihre Studie stütze sich "nicht auf überprüfbare Belege" (S. 186). Wie wir oben am Beispiel des Hox-Gens gesehen haben, scheint es sich bei diesem "Argument" um FEDUCCIAs generelle Methode zu handeln, Fakten zu ignorieren, die ihm nicht passen.

LINGHAM-SOLIAR argumentierte, dass mehrere lange, gerade, nicht spitz zulaufende "Fasern", die die Fossilien des Theropoden Sinosauropteryx umgeben, als Kollagenfasern identifiziert werden könnten. Die Merkmale, die er im Sinn hatte, entpuppten sich als Präparationsspuren von Werkzeugen, wie die hier gezeigten. Maßstab: 10 mm. Bilder: SMITHWICK et al. (2017).
Die Quintessenz zu diesem Thema ist, dass die von FEDUCCIA propagierte Kollagen-Hypothese auf oberflächlicher Ähnlichkeit und Spekulation beruht. Die bisher veröffentlichten taphonomischen und mikrostrukturellen Analysen stützen FEDUCCIAs naive Behauptung nicht, man finde "Kollagen, überall Kollagen!" (S. 190).9)
FEDUCCIAs argumentative Unbeweglichkeit in der Debatte zum Ursprung des Vogelflugs
Auch FEDUCCIAs Kapitel über WAIR10) ist voller Strohmannargumente und subjektiver Meinungen. So lautet eine weitere von FEDUCCIAs unverrückbar festbetonierten Positionen, dass der Vogelflug von den Bäumen aus entstanden sei, so dass jedes Modell, das die Entstehung des Vogelflugs vom Boden aus postuliert, zu verwerfen sei.
WAIR wurde von Ken DIAL und Kollegen um das Jahr 2003 veröffentlicht. Das Modell stand eine Zeit lang im Rampenlicht, da es eine gute Erklärung dafür zu liefern schien, wie Maniraptoren ihre ersten Flugversuche unternommen haben könnten. Die Dinge gerieten ins Wanken, als klar wurde, wie sehr ein Vogel auf das Fliegen spezialisiert sein muss, damit WAIR funktioniert. Heute ist WAIR als Erklärung für die Anfänge des Vogelflugs nicht mehr sehr beliebt. Aber FEDUCCIA kennt uns wohl besser als wir uns selbst und besteht darauf, dass WAIR noch immer eine vitale Theorie sei.
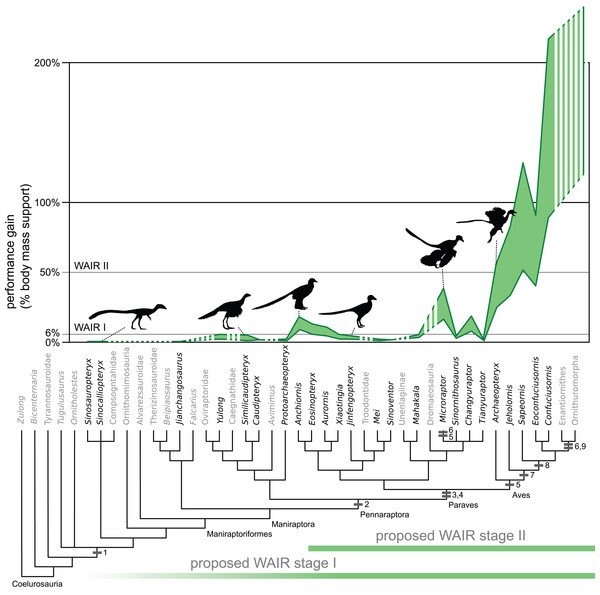
DECECCHI et al. (2016) zeigten, dass zumindest einige Maniraptoren, die keine Vögel sind, nicht die richtige Kombination anatomischer Merkmale aufweisen, um von WAIR zu profitieren, wie ursprünglich angenommen wurde. Bild: DECECCHI et al. (2016).
FEDUCCIA behauptet des Weiteren, Modelle, bei denen die Flügel für den Antrieb vom Boden aus sorgen, seien "gekommen und gegangen" (S. 224). Auch das ist nicht korrekt, da solche Ansichten in moderne Hypothesen über den Ursprung des Fluges eingeflossen sind (DECECCHI et al. 2016, NAISH & BARRETT 2018, LARSSON et al. 2020).11) Des Weiteren schreibt FEDUCCIA, dass "kein Wirbeltier den Flug vom Boden aus [entwickelt hat]" (S. 225; Kursivschrift vom Autor). Dies erklärt vielleicht, warum sich Vögel so sehr von anderen flugfähigen Wirbeltiergruppen unterscheiden, die allesamt Hintergliedmaßen haben, die in eine Flügelmembran integriert sind.
Eine Behauptung, die sich wie ein roter Faden durch das Buch zieht, ist die, dass die Wissenschaft früher oder später die von FEDUCCIA vertretene Meinung bestätigt habe. Dabei vergisst er anzumerken, dass er ausgewählte Argumente und Schlussfolgerungen, die seiner Sichtweise entsprechen, so präsentiert, als sei das "letzte Wort" zu dem jeweiligen Thema bereits gesprochen. Angesichts der an anderer Stelle des Buches von FEDUCCIA selbst getroffenen Aussage, dass die phylogenetischen Beziehungen zwischen archaischen Vögeln und vogelähnlichen Maniraptoren schwer zu entwirren seien, ist dies schwer nachvollziehbar.
Betrachten wir beispielsweise FEDUCCIAs Kritik an den Arbeiten über die taxonomische Einordnung des in Rumänien entdeckten Theropoden Balaur. FEDUCCIA lehnt Studien ab, die diesen als Dromaeosauriden einstufen, hebt aber jene eine Studie von CAU et al. hervor (2015), die zeige, dass er "kaum mehr als ein sekundär flugunfähiger Vogel" war (S. 5). Doch die betreffende Studie (ich bin einer der Autoren) kann angesichts der (bislang noch) instabilen Einordnung von Balaur in den phylogenetischen Studien nicht als "letztes Wort" zu diesem Thema angesehen werden.
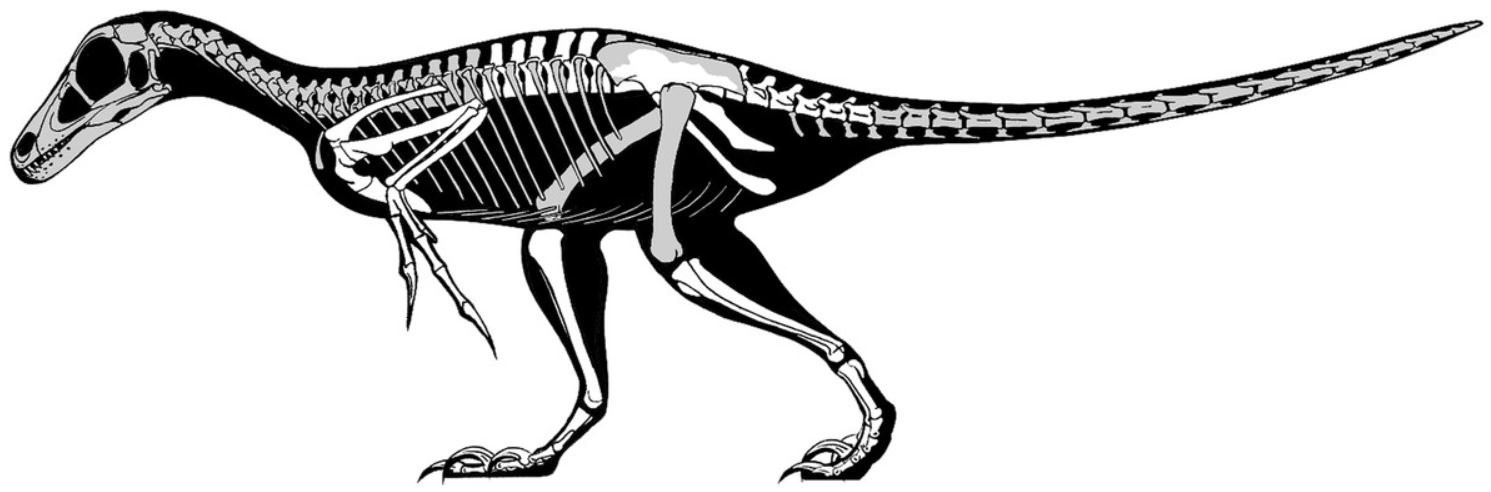
Wenn man Balaur gemäß CAU et al. (2015) als Vertreter einer urtümlichen Vogelgattung interpretiert, die den modernen Vögeln näherstand als Archaeopteryx, dann dürfte er weniger "Velociraptor-ähnlich" ausgesehen haben, als er in einigen Rekonstruktionen dargestellt wird. Aber wäre es dann fair, ihn als "kaum mehr als einen sekundär flugunfähigen Vogel" zu bezeichnen, wie FEDUCCIA vorschlägt? Bild: J. HEADDEN, mit freundlicher Genehmigung.
Physiologie
FEDUCCIAs Argumente zur Physiologie der Dinosaurier sind ebenfalls unredlich. Die allermeisten Wissenschaftler, die sich mit Nichtvogel-Dinosauriern beschäftigen, gehen davon aus, dass diese Tiere bereits eine erhöhte Stoffwechselrate besaßen. Es ist wahrscheinlich, dass die meisten oder alle Arten von Dinosauriern endotherm (warmblütig) waren. Zumindest einige Arbeiten sprechen sich für eine Meso- oder Heterothermie aus, und es könnte sein, dass die Dinosaurier verschiedene Strategien verwendeten. Dies entspricht eher unserem Verständnis von realen Tieren als eine einfache dichotome Sichtweise, da wir wissen, dass Heterothermie und sogar Ektothermie bei einigen Säugetieren vorkommt. Echte Endothermie entwickelte sich bei Knorpel- und Strahlenfischen, partielle Endothermie kommt bei einigen Echsen vor. Wie bei so vielen Dingen in der Welt der Biologie gibt es auch hier Zwischentöne und Übergänge.

Heutige Großreptilien, wie Pythons und Alligatoren, sind Dinosauriern nicht besonders ähnlich. In FEDUCCIAs Schriften wird jedoch mehrfach behauptet, die Existenz solcher Tiere sei sehr wohl von Bedeutung für die Tatsache, dass es Nichtvogel-Dinosaurier gab. Bilder: Darren NAISH.
FEDUCCIA behauptet hingegen, dass Nichtvogel-Dinosaurier mit Sicherheit "kaltblütig" waren und nennt mehrere Gründe. Zum einen seien Pythons und Alligatoren trotz ihrer wechselwarmen Physiologie zu bemerkenswertem Verhalten in der Lage, was dann auch für Dinosaurier gegolten habe. Zum anderen habe während des Mesozoikums durchgehend ein globales Treibhausklima geherrscht. Er behauptet dies mit bedauernswerter Regelmäßigkeit und beschreibt die Kreidezeit mehrfach als "eintönig heiß". In der Tat waren lange Abschnitte des Mesozoikums sehr warm. Aber das galt nicht für das gesamte Mesozoikum: Dinosaurier und ihre Zeitgenossen lebten, als das Klima gemäßigt, kühl oder sogar kalt war (CAVALHEIRO et al. 2021, WANG et al. 2023).
Die Ablagerungen der Unterkreide aus der Provinz Liaoning, berühmt für ihre kleinen Theropoden und Pterosaurier sowie für zahlreiche gefiederte Nichtvogel-Maniraptoren und archaische Vögel, scheinen in kühlen, alpinen Lebensräumen entstanden zu sein, um nur ein Beispiel zu nennen.
FEDUCCIA erwähnt Ringmuster in Dinosaurierknochen, die er als "ektotherme Wachstumsringe" deutet - so als ob sie ein eindeutiges Zeichen für eine kaltblütige Physiologie wären. Dabei ist die alte Schwarz-Weiß-Sicht auf solche Strukturen längst als falsch entlarvt.12) Kurzum, FEDUCCIAs gesamtes Kapitel über die Physiologie ist weitschweifig und chaotisch; es endet mit etwas, das sich liest wie eine Aneinanderreihung von Gedankenfetzen aus einem zoologischen Themen-Potpourri.
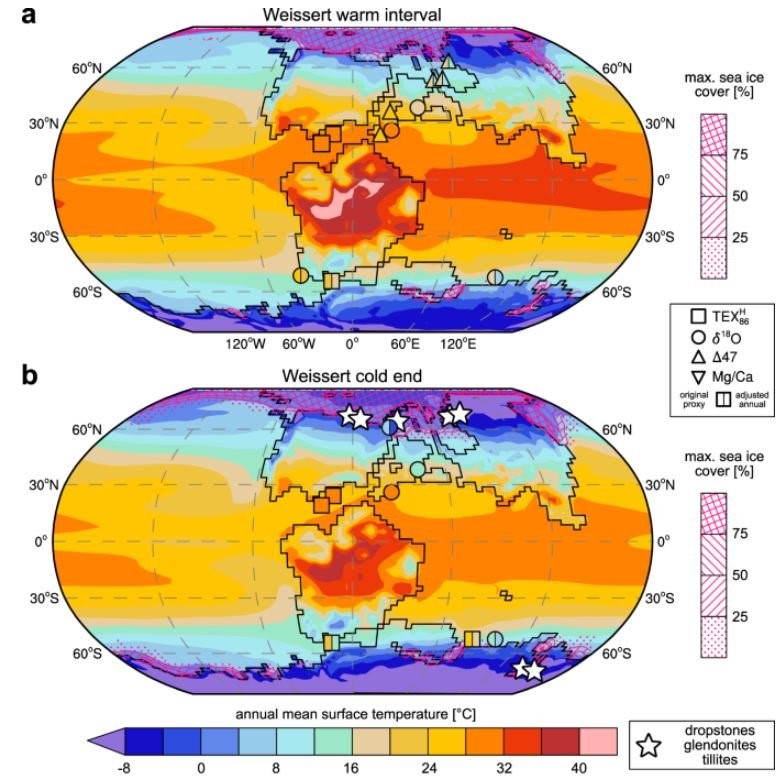
Die globalen Temperaturkarten für das Valanginium in der frühen Kreidezeit von CAVALHEIRO et al. (2021) zeigen, dass in weiten Teilen Nordamerikas und Eurasiens wohl Temperaturen von weniger als 12°C herrschten. Aus dieser Epoche sind zahlreiche Dinosaurierfossilien überliefert, die aus kühlen bis kalten Gebieten stammen. Zu beachten ist auch das für die Polarregionen angezeigte Meereis. Sicher, es gab lange Wärmeperioden im Mesozoikum. Aber die Zeiten sind vorbei, als noch angenommen wurde, dass im gesamten Mesozoikum (oder sogar noch in der Kreidezeit) ein Treibhausklima geherrscht habe. Bild: CAVALHEIRO et al. (2021).
Um Greg PAULs Willen
Ein umfangreicher Teil von RTBAD widmet sich der Idee, dass die Maniraptoren verkannte Vögel seien und auch nicht näher mit anderen Theropoden verwandt. Stattdessen soll es sich um eine Abstammungslinie handeln, die aus "vogelähnlichen Thecodonten" der Trias hervorgegangen sei. Nun argumentierte FEDUCCIA jahrzehntelang, dass Nichtvogel-Dinosaurier wie Deinonychus, Velociraptor und Oviraptor keine nennenswerte Ähnlichkeit mit archaischen Vögeln zeigen würden. Seitdem bei diesen Tieren, wie vorhergesagt, Federn entdeckt wurden, vertritt FEDUCCIA heute hingegen die genau gegenteilige Meinung - nämlich, dass diese Dinosaurier und ihre Verwandten Vögel seien... aber irgendwie trotzdem keine Dinosaurier!
Die Idee, dass Maniraptoren wie Velociraptor die flugunfähigen Nachfahren von Vögeln sein könnten, die dem Archaeopteryx ähnelten, stammt von Greg PAUL, der sie in seinem 2002 erschienenen Buch Dinosaurier der Lüfte am überzeugendsten darlegte. Kombiniert man die Belege für das Vorhandensein von Federn bei Maniraptoren mit FEDUCCIAs hartnäckigem Insistieren darauf, dass die Federn den Vogel ausmachen, wird klar, warum sich FEDUCCIA bei PAUL ankuschelt und eine verkorkste Version seiner Hypothese "alle Maniraptoren sind Vögel" übernahm. (Verkorkst, weil PAUL im Gegensatz zu FEDUCCIA Maniraptoren nach wie vor als Coelurosaurier und somit als Theropoden-Dinosaurier betrachtet). Angesichts der völligen Verachtung, die FEDUCCIA in der Vergangenheit PAUL gegenüber zum Ausdruck brachte, ist das wirklich amüsant.

Der verstorbene Larry MARTIN prägte den Begriff "Paulian School of Bird Origins" und signalisierte Zustimmung zu PAULs Hypothese, wonach die Maniraptoren sekundär flugunfähige Vögel seien. In RTBAD wird PAULs Buch Dinosaurier der Lüfte indes behandelt wie ein heiliger Text.
Um Maniraptoren von anderen Coelurosauriern und Theropoden abzugrenzen, muss man die Existenz von Maniraptoren-ähnlichen Nicht-Maniraptoren erklären. FEDUCCIA versucht dies, indem er behauptet, dass alle nicht-maniraptorischen Theropoden "kurze, stummelartige Hände" (S. 132, S. 163) hatten und ihre Vordergliedmaßen so reduziert waren, dass die Entwicklung langer Gliedmaßen dem DOLLO'schen Gesetz widersprechen würde. Er behauptet dies explizit und widmet dieser Behauptung ein ganzes Kapitel (Kapitel 10: "You Can't Go Home Again; DOLLO's Law", S. 129-135).
Tatsächlich aber haben jene Coelurosaurier, die keine Maniraptoren sind (dazu gehören die Ornithomimosaurier, Tyrannosauroiden und Compsognathiden), relativ lange, schlanke Hände und Gliedmaßen, die bereits auf einen Trend zur Verlängerung der Vordergliedmaßen innerhalb dieser Gruppe hinweisen. Wenn man ehrlich ist, sehen sie so aus wie solche, von denen man annehmen würde, dass sie die Vorfahren der Maniraptoren sind. Wir wissen, dass FEDUCCIA diese Tiere kennt, da er sie hier und da in seinen Schriften erwähnt (auch in RTBAD). Folglich kann seine Argumentation hier nicht von Naivität zeugen, sondern nur von intellektueller Unredlichkeit.
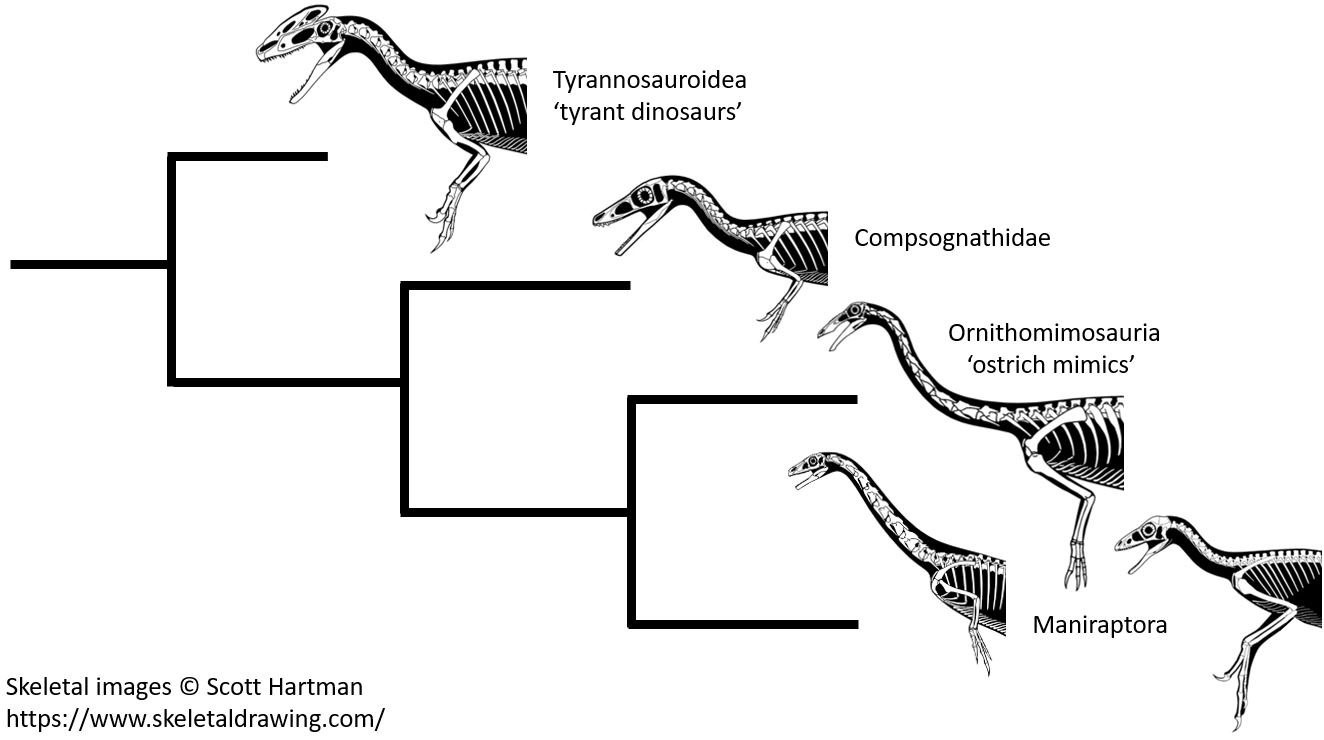
Entgegen FEDUCCIA zeigt sich bei den Theropoden, die auch die Maniraptoren umfassen, kein Trend zur Verkleinerung der Vordergliedmaßen. Wie das hier gezeigte Kladogramm andeutet, hatten sowohl Vertreter der Tyrannosauroidea (wie beispielsweise Guanlong, ein Vorfahr des Tyrannosaurus rex [MN & AB]) als auch die Ornithomimosaurier genauso wie die Maniraptoren relativ lange Arme im Vergleich zu Theropoden, die den Vögeln weniger ähneln. Ursprüngliche Maniraptoren (wie die Therizinosaurier) hatten dagegen proportional kürzere Arme als die vogelähnlicheren Maniraptoren. Bilder: © Scott HARTMAN, mit freundlicher Genehmigung.
Zu den neuesten mesozoischen Maniraptoren-Gruppen gehören die ungewöhnlichen Scansoriopterygiden, und FEDUCCIA widmet dieser Gruppe einige Aufmerksamkeit, wobei er sie oft (aber nicht immer) fälschlicherweise als "Scansoriopteriden" [sic!] bezeichnet. Scansoriopterygiden sind faszinierende Tiere; sie vereinigen eine geringe Körpergröße mit einem kurzschnäuzigen Schädel, wahrscheinlichen Kletteranpassungen an den Hinterbeinen, mit langfingrigen Vorderbeinen, die (bei einigen Taxa, nicht bei allen) mit Flugmembranen ausgestattet sind. Sie sehen aus wie Vogelvorfahren, sofern man unbedingt will (man beachte die Wortwahl!), dass sich die Vögel aus kleinen Baumkletterern entwickelt haben. So kann es nicht überraschen, dass sich FEDUCCIA für sie interessiert.
Das Problem für FEDUCCIA ist nun, dass die Scansoriopterygiden zu den Theropoden aus der Gruppe der Maniraptoren zählen; sie sind entweder mit den Oviraptorosauriern verwandt oder gehören zur Gruppe der Dromaeosauriden. Daher bedient sich FEDUCCIA eines argumentativen Tricks, der zuvor bereits von Stephen CZERKAS und Chongxi YUAN sowie von CZERKAS und FEDUCCIA Verwendung fand. Er besteht darin zu sagen, dass die Scansoriopterygiden "keine hervorstechenden Dinosauriermerkmale" (S. 243) und "keine eindeutigen Theropoden-Merkmale" (S. 247) besäßen. Das impliziert, dass die Scansoriopterygiden zwar für die Abstammung von Vögeln relevant seien, dabei seien sie aber keine Theropoden bzw. Dinosaurier gewesen.

Die Scansoriopterygiden sind wirklich faszinierende Tiere. Alle Vertreter sind klein, sie erreichen nur Sperlings- bis Taubengröße. Hier sehen wir eine Lebendrekonstruktion von Epidendrosaurus, der vermutlich mit Scansoriopteryx identisch ist. Bild: Matthew Martyniuk (Dinoguy2), Epidendrosauruswiki1, CC BY 3.0.
{kind=link}
Auch hier konfrontiert uns FEDUCCIA wieder mit seinen subjektiven Ansichten anstatt mit dem, was tatsächlich empirisch belegt ist. Die Behauptung, dass sich die Scansoriopterygiden grundlegend von Theropoden und anderen Dinosauriern unterscheiden würden, ist schlicht falsch, da ihre Skelette mit Theropoden-Merkmalen förmlich übersät sind.
Die Merkmale der Scansoriopterygiden, die FEDUCCIA als "andersartig" ansieht (wie etwa ein besonders langer dritter Finger, ein nach vorn gerichtetes Schambein, eine teils geschlossene Hüftpfanne und die weit auseinanderstehenden Darmbeine) stellen kein Problem für ihre Theropoden-Identität dar. Es ist nur so, dass die Scansoriopterygiden sonderbare Anpassungen zeigen, wobei sich ohnehin die meisten dieser Merkmale bei anderen Theropoden ebenfalls entwickelt haben.
"Konsilienz" statt Konsens
Das letzte Kapitel in RTBAD vertritt den Anspruch, "eine Suche nach «Konsilienz» statt nach Konsens" vorzunehmen. FEDUCCIA hebt nochmals die Unumstößlichkeit seiner Position hervor. Gleichzeitig rügt er all jene, die für die Abstammung der Vögel von den Dinosauriern eintreten, wegen ihrer scheinbar abwegigen Behauptungen, die keiner angemessen skeptischen Prüfung unterzogen worden seien.
Aber Moment mal: Die gefiederten Maniraptoren sind eindeutig mit den Compsognathiden, Tyrannosauroiden usw. verwandt, diese wiederum eindeutig mit den Allosauroiden usw. Die "außergewöhnliche Behauptung", die skeptisch geprüft werden sollte, ist demnach nicht die Theropoden-Abstammung der Vögel, sondern die These, dass die Maniraptoren von vierbeinigen, baumbewohnenden Nicht-Dinosaurier-Reptilien abstammen. Die Prüfung dieser These hat auch längst stattgefunden, wobei sich gezeigt hat, dass sie nicht nur unzureichend begründet ist, sondern im Widerspruch zu allen uns vorliegenden Daten steht.
Eine weitere Ironie ergibt sich aus FEDUCCIAs Mantra, dass die Federn den Vogel bestimmen: "Wenn es Federn und Flügel zum Fliegen hat, ist es ein Vogel" (S. 312). Da Lebewesen aber nun einmal aus einem Evolutionsprozess hervorgehen: Wie ist es angesichts dessen dann um Tiere bestellt, die über Protoflügel verfügten (wie etwa der Ornithomimosaurier Harpymimus, siehe ZELENITSKY et al. 2012 [MN & AB])? Wie steht es um Arten, die verschiedene Entwicklungsstadien der Federn zeigen?
Das "Vögel-sind-Dinosaurier"-Paradigma sagte solche Tiere vorher - und tatsächlich haben wir sie in Form von nicht-maniraptorischen Coelurosauriern auch gefunden. FEDUCCIA indes erinnert uns daran, dass sie in dem "Vögel-sind-keine-Dinosaurier"-Paradigma, das er uns verkaufen möchte, nicht existieren dürfen. Das ist nicht nur unwissenschaftlich, es ist anti-wissenschaftlich: Man sollte Hypothesen und Theorien bevorzugen, die mit den Daten in Einklang stehen, nicht umgekehrt!
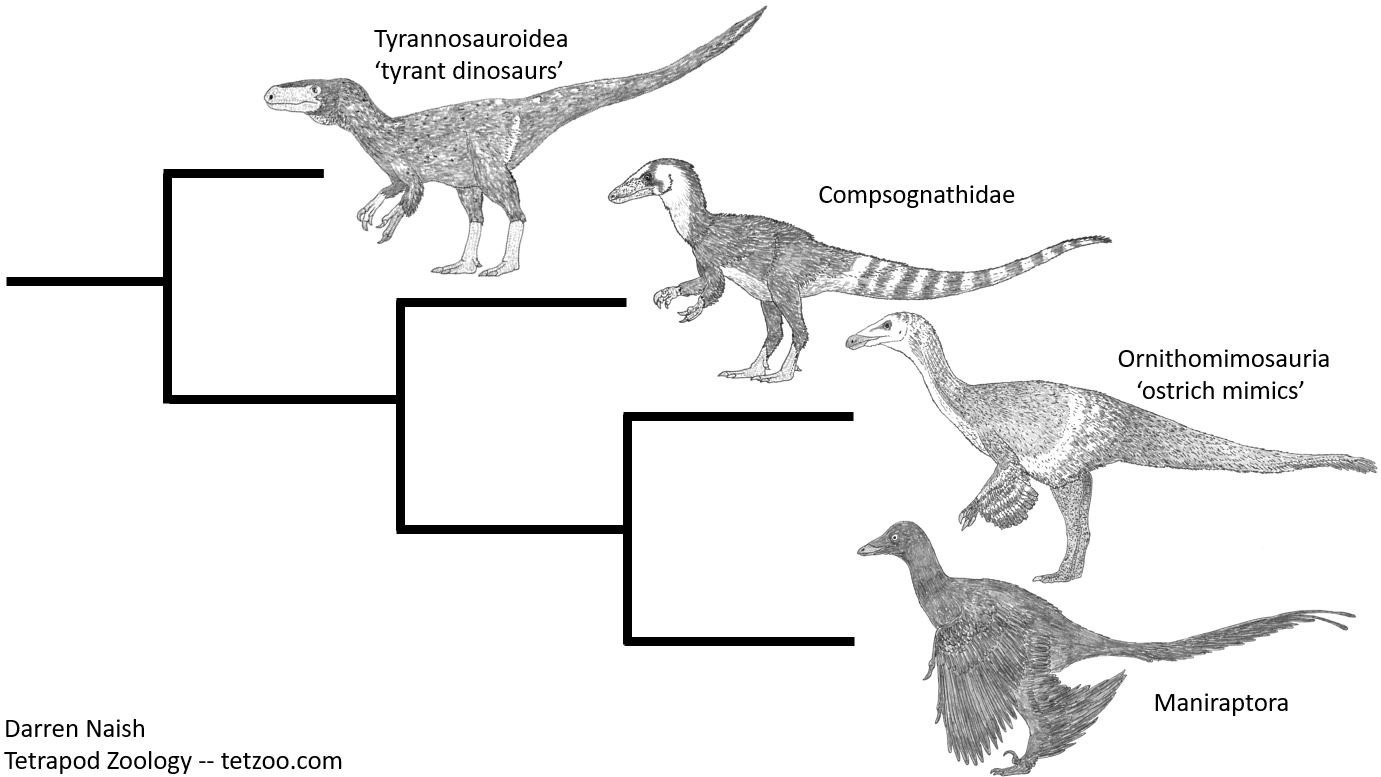
Die Maniraptoren, zu denen auch die Vögel gehören, existieren nicht in einem phylogenetischen Vakuum. Das heißt, sie sind nicht grundlegend anders als die übrigen Theropoden und teilen zahlreiche anatomische Merkmale mit Ornithomimosauriern, Compsognathiden und Tyrannosauroiden. Die Gemeinsamkeiten sind so zahlreich, dass jede phylogenetische Hypothese, die den Maniraptoren eine Bedeutung in der Vogelevolution zumisst, auch diese anderen Theropoden-Gruppen "mitziehen" muss. Diese Abbildung zeigt eine stark vereinfachte Verwandtschaftshypothese für die genannten Gruppen. Bild: Darren NAISH.
Das alles führt mich zu meinem letzten Punkt. Wie offensichtlich geworden sein dürfte, hat mir dieses Buch nicht gefallen. Ich war ständig frustriert von der Rosinenpickerei des Autors, was den selektiven Umgang mit Daten und Zitaten anbelangt.
FEDUCCIAs Versuch, aktuelle Ansichten über die Welt des Mesozoikums zu diskreditieren, sorgte zusehends für Verdruss. Nicht zuletzt war ich genervt von der hartnäckigen Kampagne zur Verbreitung seiner Sicht, dass Wissenschaftler, die sich mit Dinosauriern beschäftigen, verzweifelte Selbstdarsteller und Größenwahnsinnige seien, die in einer Art Sekte agieren. Wie erwähnt sind seine rhetorischen Stilmittel des Whataboutismus, der Strohmann-Argumente und der Wiederholung von Falschbehauptungen allgegenwärtig und verwandeln RTBAD in ein frustrierendes, voreingenommenes und intellektuell problematisches Propagandastück.
Trotz alledem kann ich als jemand, der von konträren und unorthodoxen Ansichten in der Wissenschaft fasziniert ist, nur sagen, dass ich froh bin, dass es dieses Buch gibt. Ja, es ist eine gewaltige Ironie, dass ich mich darüber freue, dass dieses Buch erschienen ist und Alan FEDUCCIA seine konträren, intellektuell problematischen, wissenschaftlich unredlichen und handwerklich fehlerhaften Gedanken zur Evolution und Biologie der Dinosaurier und ihrer Verwandten niederschrieb.

Alan FEDUCCIA vertritt seit Jahrzehnten die Ansicht, dass er und seine Kollegen "die Vögel kennen", während diejenigen, die das "Vögel-sind-Dinosaurier"-Paradigma vertreten, dies nicht tun. Das hat nie gestimmt, und es wird mit der Zeit immer offensichtlicher. Wie dem auch sei, wir könnten FEDUCCIAs Unterstellung problemlos spiegeln und ihm vorhalten, dass er mit Sicherheit von Nichtvogel-Dinosauriern keine Ahnung hat. Bilder: Darren NAISH.
Profitieren wir von FEDUCCIAs Bemühungen, den wissenschaftlichen Konsens zu entkräften, wie einige meinen? Ich glaube, dass das Gegenteil der Fall ist.13) Doch ich gebe zu, dass ich mich am Schrägen erfreue.
Danksagung
Diese Rezension ist dank der Großzügigkeit von Martin NEUKAMM entstanden. Ich danke auch Kollegen, die den Text kommentiert und Teile davon kritisch überprüft haben, nämlich Albert CHEN, John HARSHMAN, Gerald MAYR, Mike P. TAYLOR und Mathew WEDEL.
© Darren NAISH (nur Text). Übertragung mit freundlicher Genehmigung des Autors.
Literatur
DE BAKKER, M. A. G., VAN DER VOS, W., DE JAGER, K. et al. (2021). Selection on phalanx development in the evolution of the bird wing. Molecular Biology and Evolution 38, 4222-4237.
ČAPEK, D., METSCHER, B. D. & MÜLLER, G. B. (2013). Thumbs down: a molecular‐morphogenetic approach to avian digit homology. Journal of Experimental Zoology (Molecular and Developmental Evolution) 9999B, 1-12.
CAU, A. (2018). The assembly of the avian body plan: A 160-million-year long process. Bollettino della Società Paleontologica Italiana 57, 1-25.
CAU, A., BROUGHAM, T. & NAISH, D. (2015). The phylogenetic affinities of the bizarre Late Cretaceous Romanian theropod Balaur bondoc (Dinosauria, Maniraptora): dromaeosaurid or flightless bird? PeerJ 3: e1032.
CAVALHEIRO, L., WAGNER, T., STEINIG, S. et al. (2021). Impact of global cooling on Early Cretaceous high pCO2 world during the Weissert Event. Nature Communications 12: 5411.
CHRISTIAN, A. & DZEMSKI, G. (2011). Neck posture in sauropods. In KLEIN, N., REMES, K., GEE, C. T. & SANDER. P. M. (Eds.) Biology of the Sauropod Dinosaurs (pp. 251-260). Indiana University Press, Bloomington and Indianapolis.
COOPER, A., LALUEZA-FOX, C., ANDERSON, S. et al. (2001). Complete mitochondrial genome sequences of two extinct moas clarify ratite evolution. Nature 409, 704-707.
CURRIE, P. J. & CHEN, P.-J. (2001). Anatomy of Sinosauropteryx prima from Liaoning, northeastern China. Canadian Journal of Earth Sciences 38, 1705-1727.
DECECCHI, T. A., LARSSON, H. C. E. & HABIB, M. B. (2016). The wings before the bird: an evaluation of flapping-based locomotory hypotheses in bird antecedents. PeerJ 4: e2159.
FEDUCCIA, A. (1977a). The whalebill is a stork. Nature 266, 719-720.
FEDUCCIA, A. (1977b). Hypothetical stages in the evolution of modern ducks and flamingos. Journal of Theoretical Biology 67, 715-721.
FEDUCCIA, A. (1978). Presbyornis and the evolution of ducks and flamingos. American Scientist 66, 298-304.
FEDUCCIA, A. (1996). The Origin and Evolution of Birds. Yale University Press, New Haven & London.
FEDUCCIA, A. (2002). Birds are dinosaurs: simple answer to a complex problem. The Auk 119, 1187-1201.
FEDUCCIA, A. (2012). Riddle of the Feathered Dragons: Hidden Birds of China. Yale University Press, New Haven & London.
FEDUCCIA, F. (2020). Romancing the Birds and Dinosaurs: Forays in Postmodern Paleontology. BrownWalker Press, Irvine, Boca Raton.
FIELD, D. J., BENITO, J., CHEN, A. et al. (2020). Late Cretaceous neornithine from Europe illuminates the origins of crown birds. Nature 579, 397-401.
GAUTHIER, J. A. (1986). Saurischian monophyly and the origin of birds. In PADIAN, K. (Ed.) The origin of birds and the evolution of flight (p. 1-55). California Academy of Science, San Francisco.
GODEFROIT, P., SINITSA, S. M., CINCOTTA, A. et al. (2020). Integumentary structures in Kulindadromeus zabaikalicus, a basal neornithischian dinosaur from the Jurassic of Siberia. In FOTH, C. & RAUHUT, O. W. M. (Eds.) The Evolution of Feathers (pp. 48-65). Springer Nature, Berlin.
GODEFROIT, P., SINITSA, S. M., DHOUAILLY, D. (2014). Response to Comment on 'A Jurassic ornithischian dinosaur from Siberia with both feathers and scales'. Science 346, 434.
LARSSON, H. C. E., DECECCHI, T. A. & HABIB, M. B. (2020). Navigating functional landscapes: a bird's eye view of the evolution of avialan flight. In Pittman, M. & Xu, X. (Eds.) Pennaraptoran Theropod Dinosaurs: Past Progress and New Frontiers (pp. 321-332). Bulletin of the American Museum of Natural History 440, American Museum of Natural History, New York.
LINGHAM-SOLIAR, T. (2001). The ichthyosaur integument: skin fibers, a means for a strong, flexible and smooth skin. Lethaia 34, 287-302.
LINGHAM-SOLIAR, T. & WESLEY-SMITH, J. (2008). First investigation of the collagen D-band ultrastructure in fossilized vertebrate integument. Proceedings of the Royal Society B 275, 2207-2212.
MAYR, G. (2010). Response to Lingham-Soliar: dinosaur protofeathers: pushing back the origin of feathers into the Middle Triassic? Journal of Ornithology 151, 523-524.
MAYR, G., PITTMAN, M., SAITTA, E. T. et al. (2016). Structure and homology of Psittacosaurus tail bristles. Palaeontology 59, 793-802.
MORELL, V. (1997). The origin of birds: the dinosaur debate. Audubon 99 (2), 36-45.
NAISH, D. & BARRETT, P. M. (2018). Dinosaurs: How They Lived and Evolved. The Natural History Museum, London.
NESBITT, S. J., SMITH, N. D., IRMIS, R. B. et al. (2009) A complete skeleton of a late Triassic Saurischian and the early evolution of dinosaurs. Science 326, 1530-1533.
NEUKAMM, M. & BEYER, A. (2023). Why do we know that birds are living dinosaurs?
OLSON, S. L. & FEDUCCIA, A. (1980a). Relationships and evolution of flamingos (Aves: Phoenicopteridae). Smithsonian Contributions to Zoology 316, 1-73.
OLSON, S. L. & FEDUCCIA, A. (1980b). Presbyornis and the origin of the Anseriformes (Aves: Charadriomorphae). Smithsonian Contributions to Paleobiology 323, 1-24.
PRUM, R. O. (2001). Longisquama fossil and feather morphology. Science 291, 1899-1900.
RAUHUT, O. W. M. (2003). The interrelationships and evolution of basal theropod dinosaurs. Special papers in paleontology 69, 1-213.
ROWE, T., KETCHAM, R. A., DENISON, C. et al. (2001). Forensic palaeontology: the Archaeoraptor forgery. Nature 410, 539-540.
SALINAS-SAAVEDRA, M., GONZALEZ-CABRERA, C., OSSA-FUENTES, L. et al. (2014). New developmental evidence supports a homeotic frameshift of digit identity in the evolution of the bird wing. Frontiers in Zoology 11: 33.
SERENO, P. C. (1999). The evolution of dinosaurs. Science 284, 2137-2147.
SHAHID, F., ZHAO, J., & GODEFROIT, P. (2019). Aerodynamics from cursorial running to aerial gliding for avian flight evolution. Applied Sciences 9, 649.
SMITHWICK, F. M., MAYR, G., SAITTA, E. T. et al. (2017). On the purported presence of fossilized collagen fibres in an ichthyosaur and a theropod dinosaur. Palaeontology 60, 409-422.
STEWART, T. A., LIANG, C., COTNEY, J. L. et al. (2019). Evidence against tetrapod-wide digit identities and for a limited frame shift in bird wings. Nature Communications 10: 3244.
TAYLOR, M. P., WEDEL, M. J. & NAISH, D. (2009). Head and neck posture in sauropod dinosaurs inferred from extant animals. Acta Palaeontologica Polonica 54, 213-220.
UNWIN, D. M. & BENTON, M. J. (2001). Longisquama fossil and feather morphology. Science 291, 1900-1901.
VARGAS, A. O. & FALLON, J. F. (2005). The digits of the wing of birds are 1, 2 and 3. A review. Journal of Experimental Zoology (Molecular and Developmental Evolution) 304B, 206-219.
VIDAL, D., MOCHO, P., ABERASTURI, A. et al. (2020a). High browsing skeletal adaptations in Spinophorosaurus reveal an evolutionary innovation in sauropod dinosaurs. Scientific Reports 10 (6638).
VIDAL, D., MOCHO, P., PÁRAMO, A. et al. (2020b). Ontogenetic similarities between giraffe and sauropod neck osteological mobility. PLoS ONE 15 (1): e0227537.
ZELENITSKY, D., THERRIEN, F., ERICKSON, G. M. et al. (2012) Feathered non-avian dinosaurs from North America provide insight into wing origins. Science 338, 510-514.
ZHANG, F., KEARNS, S. L., ORR, P. J. et al. (2010). Fossilized melanosomes and the colour of Cretaceous dinosaurs and birds. Nature 463, 1075-1078.
Fußnoten
[1] Ins Deutsche übertragen von Martin Neukamm & Andreas Beyer. Das Original findet sich unter https://tetzoo.com/blog/2023/10/2/alan-Feduccia-romancing-the-birds-and-dinosaurs
[2] FEDUCCIA meint damit die etablierte Klassifikationsmethode der phylogenetischen Systematik (Kladistik), welche die verwandtschaftlichen Beziehungen zwischen den Arten rekonstruiert. Phylogenie oder Phylogenese meint die Evolutionsgeschichte bestimmter Arten oder der Lebewesen allgemein. [MN & AB]
[3] Bestätigungsverzerrung ist eine Art der kognitiven Denkverzerrung, die dazu führt, dass Informationen einseitig verarbeitet und so interpretiert werden, so dass sie scheinbar die eigene (voreingenommene) Meinung bestätigen. Dabei werden Befunde, die nicht zur eigenen Meinung passen, außer Acht gelassen, für falsch oder unvollständig erklärt oder auf bestätigende Teilaspekte reduziert. [MN & AB]
[4] BRUSATTE ist einer der "Mainstream-Paläontologen" auf diesem Gebiet. [MN & AB]
[5] Die Deutung dieser Körperanhängsel als Federn hält ein Großteil der Paläontologen für eine Fehlinterpretation der Gewebsabdrücke im Gestein, die widersprechende Befunde nicht berücksichtigt (vgl. etwa PRUM 2001; UNWIN & BENTON 2001). Am ehesten dürfte es sich um verlängerte Rückenschuppen handeln. Auch die These, es handele sich um eine Anpassung an einen Gleitflug, ist reine Spekulation. FEDUCCIA tut indes so, als handele es sich um fundierte Erkenntnisse, weil die Spekulationen gut in sein "Vögel-sind-keine-Dinosaurier"-Paradigma passen (Stichwort Bestätigungsverzerrung). [MN & AB]
[6] Die Vikarianz-Hypothese besagt in diesem Zusammenhang, dass alle Laufvögel einen gemeinsamen Vorfahren hatten, der den Großkontinent Gondwana in der späten Kreidezeit besiedelte. Als der Großkontinent zerbrach, wurden sie durch Plattendrift an ihre aktuellen Positionen gebracht und entwickelten sich isoliert voneinander zu den heute vorhandenen Laufvogelarten. Neuere Analysen der genetischen Variation zwischen den Laufvögeln unterstützen dieses einfache Bild jedoch nicht (vgl. etwa COOPER et al. 2001). [MN & AB]
[7] Bei einigen Theropoden findet man die Fingerformel (I-II-III), was bedeutet, dass Daumen, Zeige- und Mittelfinger erhalten blieben, während Ring- und kleiner Finger teilweise oder ganz reduziert wurden. FEDUCCIA behauptet, die Vögel würden dagegen die Fingerformel (II-III-IV) besitzen, was gegen ihre Abstammung von Theropoden spräche. Allerdings sind die Befunde diesbezüglich nicht eindeutig, und die evolutionäre Entwicklungsbiologie kann auch erklären, warum das so ist. Näheres zu diesem Thema in NEUKAMM & BEYER (2023, S. 22-24). [MN & AB]
[8] Das ist allerdings bestenfalls ein "Argument aus Nichtwissen". Außerdem: Wenn einige Theropoden ihre Fingerzahl auf drei reduzierten, was der Fall ist, und dies die oben beschriebenen biomechanischen Konsequenzen haben sollte, dann wäre die embryologische Konsequenz zwingend eine Frameshift - unabhängig davon, ob das Ergebnis eine Funktion hat oder nicht. Ohnehin muss in der Biologie nicht alles eine Funktion haben! [MN & AB]
[9] Auch FOTH & RAUHUT (2020, S. 28) bemerken zu den alternativen Deutungen der Integument-Filamente als degradierte dermale Kollagenfasern oder andere Gewebe, die betreffenden Studien seien "…aus taphonomischen, strukturellen und methodischen Gründen kritisiert worden (etwa MAYR 2010; SMITH et al. 2015; SMITHWICK et al. 2017) und können daher nicht aufrechterhalten werden. Darüber hinaus machen die große Anzahl von Taxa, bei denen inzwischen Federn nachgewiesen wurde, sowie die große Vielfalt an Federtypen (zum Beispiel XU & GUO 2009) diese alternativen Interpretationen unhaltbar." [MN & AB, ebenso Übersetzung ins Deutsche]
[10] Das Kürzel WAIR steht für wing-assisted incline running. Dabei handelt es sich um ein Modell, wonach der Vogelflug über eine Art "Flattern", mit denen junge Vögel Steigungen und Hindernisse bewältigen können, vom Boden aus seinen Anfang genommen haben soll. [MN & AB]
[11] Ein interessantes Szenario, das die Stärken der "Boden-" und der "Baumtheorie" kombiniert und die scheinbaren Gegensätze zum Verschwinden bringt, diskutiert SHAHID et al. (2019). So hat sich gezeigt, dass Aufwinde, etwa an Bergflanken, kleine Paravianer vom Boden aus bis in die Bäume tragen können. [MN & AB]
[12] Tatsächlich existieren derlei Wachstumsringe auch bei Säugetieren; außerdem wuchsen Dinosaurier sehr rasch, und dafür brauchen sie eine hohe Stoffwechselrate, vgl. https://www.deutschlandfunk.de/jahresringe-bei-saeugetieren-100.html [MN & AB]
[13] Tatsächlich dient, wie nicht anders zu erwarten, das Werk bereits als Erbauungslektüre der Evolutionsleugner. Daher ist es unerlässlich, derlei demagogischen Meisterwerken mit eloquenter Sachkritik und wissenschaftlicher Aufklärungsarbeit "den Wind aus den Segeln" zu nehmen. Wer sich vertiefend mit dieser Thematik beschäftigen möchte, dem sei das Review-Paper NEUKAMM & BEYER (2023) empfohlen. [MN & AB]


